DZIAŁANIE PRĄDÓW ELEKTRYCZNYCH NA USTRÓJ
Dodatnie jak i ujemne skutki, jakie powoduje prąd elektryczny przepływając przez organizm ludzki, zawsze były w kręgu zainteresowania człowieka. Prace nad wpływem prądu na organizm zaczęły się zaraz po zbudowaniu przez Alessandro Volta pierwszego ogniwa galwanicznego w 1800 roku. Sam Volta badał wpływ bodźców elektrycznych na zmysł wzroku i smaku. (Najbardziej chyba powszechnie znanym, ujemnym skutkiem działania prądu jest porażenie elektryczne). Dopiero jednak w ostatnich kilkudziesięciu latach wiedza na ten temat poszerzyła się znacznie. Badania dokonywane w wielu krajach, także w Polsce, odpowiednio ją usystematyzowały.
Niniejszy artykuł ma na celu przybliżenie czytelnikowi ogólne działanie prądu na komórkę ludzką i zasygnalizowanie obszerności tematu, nie wgłębiając się w tematykę prądów selektywnych, opisywanych w innych publikacjach dostępnych na rynku.
Rodzaje prądów elektrycznych.
W fizjologii i w medycynie stosuje, się w różnych celach prądy elektryczne różnego rodzaju np.:
Prąd stały (galwaniczny) - jest to prąd jednokierunkowy, przeważnie niskiego napięcia (do 80 woltów)
Prąd stały przerywany (galwaniczny przerywany) - jest to prąd jednokierunkowy niskiego napięcia, przerywany systematycznie z częstotliwością od 30 razy na minutę do 120 razy na minutę (kształt impulsu prostokątny).
Prąd stały przerywany o regulowanym kształcie impulsu ( dla prądów selektywnych jest to kształt trapezu) o częstotliwościach rzędu kilku Hz.
Prąd zmienny sinusoidalny (sinus-farad) -jest to prąd zmienny o częstotliwości 50-90 Hz (poszczególne impulsy trwają około 0,02-0,01 s); prąd taki otrzymuje się z sieci oświetleniowej, a w badaniach fizjologicznych otrzymuję się z układu silnik-prądnica, albo też z generatora prądowego.
Prąd faradyczny normalny jest to prąd zmienny niskiego napięcia, asymetryczny przerywany o częstotliwości rzędu kilkuset Hz (poszczególne impulsy trwają około 0,001 s); prąd taki otrzymuje się najczęściej z cewki indukcyjnej Ruhmkorfa
Prąd faradyczny modulowany - jest to prąd faradyczny o zmienianej stopniowo amplitudzie, a czasem o zmienianej częstotliwości impulsów.
Prąd diatermiczny - jest to prąd zmienny o wielkiej częstotliwości rzędu 500 kHz do 100 MHz, tzn. o długości fali od 600 metrów do 3 metrów; najczęściej stosuje się prąd diatermiczny długofalowy (długości fali około 300 metrów) i prąd diatermiczny krótkofalowy (długości fali od 3 do 12 metrów)
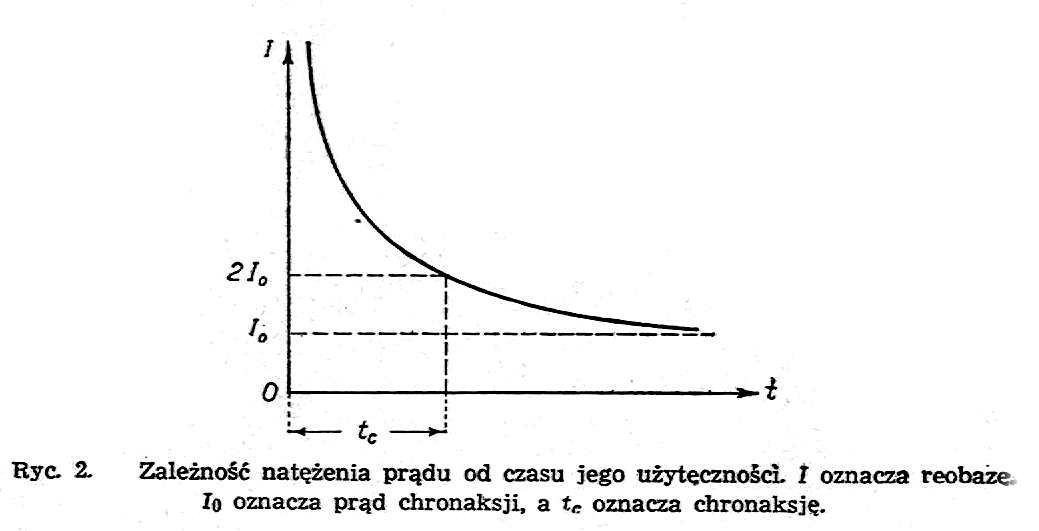
Najmniejszy czas działania prądu- stałego; potrzebny do spowodowania reakcji fizjologicznej, nazywamy czasem użytecznym. Czas użyteczny służy za miarę pobudliwości: im czas jest krótszy, tym pobudliwość jest większa.
Ponieważ bezpośrednie badania progu pobudliwości dają wyniki trochę rozbieżne, przeto dla określenia pobudliwości nerwu i mięśni zostało wprowadzone w fizjologii i w medycynie pojęcie tzw. chronaksji Określa się je mianowicie w ten sposób, że wyznacza się najpierw minimalna wartość prądu stałego, dającą pobudzenie (wartość ta nosi nazwę reobazy),a następnie podwaja się ją i dla tej podwojonej wartości prądu wyznacza się jego czas użyteczny. Ten czas nosi nazwę chronaksji. Na ryc.2 pokazany jest wykres zależności wartości natężenia prądu stałego od jego czasu użyteczności; I0 oznacza reobazę, 2I0 - prąd chronaksji, tc - chronaksję.
Prądy wielkiej częstotliwości wykazują nawet przy dużym napięciu i dużej gęstości słabe działanie elektrochemiczne, a to z powodu częstej zmiany kierunku napięcia; jony elektrolityczne mają dużą masę, a więc i dużą bezwładność; zyskują wobec tego małe przyspieszenie i nie mogą podczas trwania napięcia jednego kierunku przesunąć się dostatecznie daleko, żeby spowodować wyraźną zmianę koncentracji. Dlatego też; końce wtórnego uzwojenia transformatora Tesli można trzymać w rękach bez szkody dla organizmu, chociaż amperomierz, włączony szeregowo w obwód z ciałem, wskazuje przepływ prądu rzędu kilku miliamperów i chociaż żarówki włączone do tego obwodu świecą jasno.
Dawniej przypuszczano, że efekt ten pochodzi stąd, że prądy wielkiej częstotliwości przepływają w tym przypadku tylko po powierzchni człowieka (tzw. efekt naskórkowy). Pogląd ten jednak nie jest słuszny, gdyż efekt tego rodzaju mogą dawać tylko bardzo dobre przewodniki, jak metale, ale nie takie, jakimi są organizmy.
Mechanizm działania prądów elektrycznych. Napięcia i prądy czynnościowe.
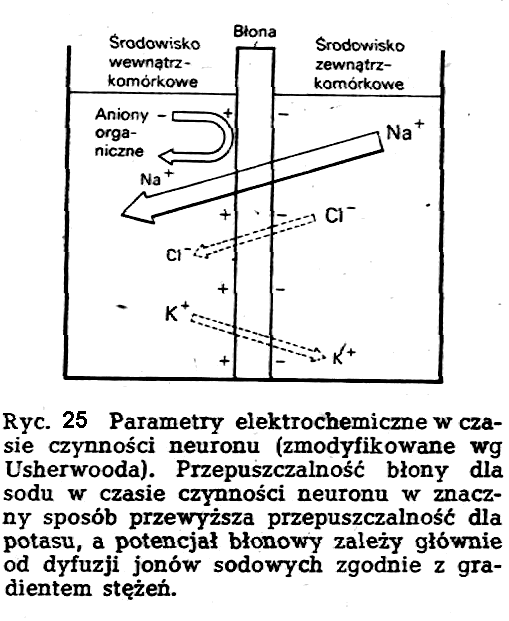
Tkanki ciała ludzkiego składają się z substancji elektrolitycznych o oporze właściwym rzędu kilkuset Wm. Tkanki zawierają jony różnych soli, przeważnie jony Na (dodatnie) i jony Cl (ujemne) - w ilości około 6 g na 1l, a poza tym zawierają już w znacznie mniejszej ilości jony: K, Ca, Mg, P i inne.
W tkankach rozłożone są równomiernie komórki zamknięte półprzenikliwą błoną, której opór elektryczny jest znacznie większy od oporu otaczającej je substancji. Wewnątrz komórek znajduje się substancja elektrolityczna o podobnych właściwościach elektrycznych, jak substancja zewnętrzna. Kształt komórek można pominąć początkowo przy ogólnych rozważaniach, podobnie jak i strukturę wewnętrzną komórki. Opór elektryczny pojedynczej komórki jest spowodowany prawie wyłącznie przez opór błony komórkowej. Opór elektryczny tkanki dla prądu stałego jest określony głównie przez opór elektryczny pojedynczej błony komórkowej i przez liczbę komórek przypadających na 1 cm3 tkanki. Wartość oporu elektrycznego tkanki dla prądu stałego nie ma na ogół większego znaczenia praktycznego, stanowi ona natomiast ważną podstawę dla badania mechanizmu przewodzenia prądów zmiennych.
Pod wpływem przyłożonego napięcia zewnętrznego jony przesuwają się w polu elektrycznym wewnątrz komórek i w przestrzeniach międzykomórkowych, wskutek czego w poszczególnych częściach tkanki przepływa prąd elektryczny i powstaje zmiana koncentracji jonów, wywołująca polaryzację elektrolityczną i miejscowe różnice potencjałów elektrycznych. Zjawiska te mają duże znaczenie fizjologiczne, a w szczególności mogą wywołać podrażnienia tkanki, jeżeli tylko podnieta elektryczna przekroczy pewną charakterystyczną dla danej tkanki wartość, tzw. próg pobudliwości.
Dla komórek nerwowych i mięśniowych, które mają kształt walcowy, zjawisko przewodnictwa elektrycznego na poszczególnych odcinkach przebiega podobnie jak dla kabli elektrycznych. Spadek potencjału przy przepływie prądu wzdłuż komórki następuje wtedy zgodnie z prawem Ohma, a spadek potencjału w poprzek komórki można pominąć przy rozpatrywaniu oporu całej tkanki.
Inaczej przedstawia się sprawa przewodzenia przez komórki i tkanki prądu zmiennego, mianowicie w tym przypadku należy uwzględnić opór pojemnościowy i opór samoindukcyjny komórek, podobnie jak przy obliczaniu zawady elektrycznej . Opory te zależą od częstotliwości kołowej co i tylko w niektórych przypadkach, kiedy pojemność komórki i jej samoindukcja są tak dobrane, że opór zawady znika, zagadnienie bardzo się upraszcza i opór pozorny komórki zbliża się do oporu omowego.
Zmiany koncentracji jonów są na ogół proporcjonalne do gęstości prądu (tzn. do natężenia prądu przypadającego na jednostkę powierzchni ciała), dlatego największe zmiany fizjologiczne zachodzą w tych miejscach, w których elektrody przyłożone do ciała mają powierzchnie najmniejszą (przy tym samym natężeniu prądu).
Koncentracji jonów przeciwdziała zjawisko dyfuzji jonów, dzięki któremu jony dążą do takiego położenia, przy którym rozkład ich jest najbardziej równomierny
Zjawisko dyfuzji zachodzi w tym większym stopniu, im większe są różnice koncentracji jonów i wymaga pewnego czasu dla wyrównania rozkładu jonów; z tego względu prądy krótkotrwałe (przy
których dyfuzja jonów gra rolę znikomą) wykazują znacznie silniejsze działanie drażniące niż prądy długotrwałe, przeprowadzające przez ciało te samą ilość naboju elektrycznego, tym się tłumaczy np. fakt, że prądy otwarcia w induktorze działają pod względem fizjologicznym dużo silniej niż prądy zamknięcia.
Działanie elektropatologiczne prądów elektrycznych
Najbardziej niebezpieczne dla życia są prądy zmienne o małej częstotliwości, rzędu 30-150 okresów na sekundę. Górna granica bezpieczeństwa wynosi dla nich około 200 woltów napięcia; dla prądów stałych granica ta przesuwa się do 500 woltów. Jednak przy nieszczęśliwym zbiegu okoliczności mogą zajść przypadki śmiertelne nawet przy 60 woltach prądu zmiennego i 250 woltach prądu stałego i dlatego w technice przyjmuje się jako graniczną wartość bezpieczeństwa napięcie 42 woltów prądu zmiennego.
Wypadki porażenia prądem mogą się zdarzyć np. przy dotykaniu przewodów mokrymi rękami, szczególnie w wannie (jeśli ta łączy się przypadkowo z jednym biegunem elektrycznym, przy dotknięciu ręką drugiego bieguna itp.}. Granice bezpieczeństwa są dla różnych osób różne, poza tym nagłe, nieoczekiwane uderzenia elektryczne działają dużo silniej niż uderzenia spodziewane.
W większości wypadków śmierć następuje prawdopodobnie przez zatrzymanie czynności serca wskutek drżeń włókienkowych. Przy dłuższym działaniu (ponad 2 minuty) następuje uduszenie wskutek ogólnego skurczu tężcowego mięśni. Często jednak można jeszcze uratować porażonego prądem przez zastosowanie sztucznego oddychania.
Prądy wielkiej częstotliwości nie są niebezpieczne nawet przy napięciu 100000 woltów i więcej. Działanie fizjologiczne jest w przybliżeniu odwrotnie proporcjonalne do pierwiastka kwadratowego z częstotliwości (w pewnym zakresie częstotliwości).
Do ujemnych skutków działania prądu należy też rozmnażanie się komórek rakowych w organizmie, dlatego osoby będące na chemioterapii nie powinny stosować prądów w leczeniu uzupełniającym.
Opór elektryczny ciała ludzkiego dla prądu stałego.
Mechanizm tych zjawisk jest na ogół bardzo różny w zależności od rodzaju nośników ładunku elektrycznego w danej substancji (elektronów swobodnych, jonów) jak również od rodzaju i wielkości przyłożonej siły elektromotorycznej (różnicy potencjałów), od temperatury itd.
W tkankach żywego organizmu pod wpływem przyłożonego napięcia zewnętrznego będą występowały najczęściej zjawiska typu elektrochemicznego ze względu na to, że, tkanki składają się w dużej mierze z różnego rodzaju elektrolitów, poprzedzielanych nieprzewodzącymi elektrycznie błonami. Zresztą o przewodnictwie elektrycznym tkanek decyduje najczęściej ilość zawartej w nich wody. Niezależnie jednak od tego mogą wchodzić w grę jeszcze inne zjawiska natury elektrycznej, które należy wziąć pod uwagę w poszczególnych przypadkach.
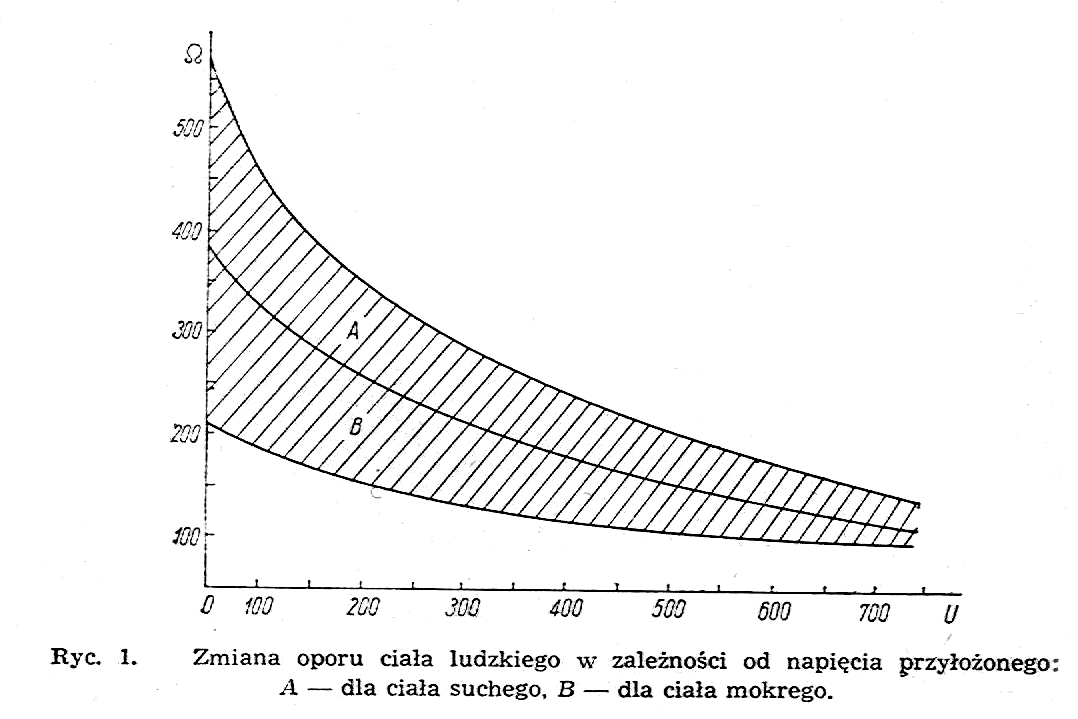
Przy pomiarach prądem stałym otrzymuje się dla całego ciała ludzkiego przy doprowadzeniu różnicy potencjałów do rąk jak ma to miejsce w przypadku stosowania prądów selektywnych, ewentualnie do jednej ręki i do przeciwległej nogi, opór elektryczny wartości rzędu paru tysięcy omów. Na rys.1 pokazana jest zależność oporu elektrycznego dla ciała ludzkiego od przyłożonego napięcia (według Freiberga), przy czym wartości oporu mogą się zmieniać w granicach pomiędzy górną i dolną krzywą w obszarze zakreskowanym w zależności od wilgotności ciała.
Opór właściwy (r) tkanki jest funkcją kilku różnych parametrów takich, jak: opór właściwy płynu zewnatrzkomórkowego (r1), opór właściwy zawieszonych w niej komórek (r2), względna objętość komórek (j), opór właściwy plazmy (r2), opór właściwy błony komórkowej (r4), promień komórki (a), i czynnika geometrycznego (f) określającego kształt komórki.
Z rozważań teoretycznych i doświadczalnych nad różnymi rodzajami zawiesin w roztworach elektrolitycznych ustalono następującą zależność dla r:
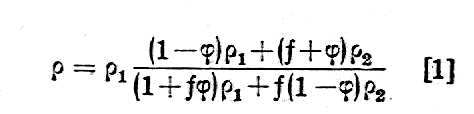
Przy tym r2 = r3 + r4/a czynnik j=vK/V określa stosunek objętości komurek (vK) do całkowitej objętości tkanki (V) a czynnik gemetryczny f przyjmuje wartości:1.5 dla komórek kulistych.
Wartości liczbowe poszczególnych oporów właściwych są rzędu: (r1~102Wm, r2~104Wm, r3~1Wm, r4~0,1Wm(przy a = 10-5m). Jak łatwo zauważyć ze wzoru [1], podstawiając różne wartości na r2, ich wpływ na wartość r1 jest stosunkowo nieduży (rzędu paru procent) tak, że można napisać wzór [1] w postaci uproszczonej
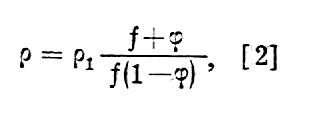
W poniższej tebeli podane są wartości liczbowe oporu właściwego różnych tkanek.
|
Tkanka |
Opór właściwy Ohm |
|
Płyn mózgowo- rdzeńiowy |
0 |
|
Surowica krwi |
0 |
|
Mięśnie |
1, 52 |
|
Krew |
0 |
|
Wątroba |
0 |
|
Mózg |
25 |
|
Tkanka tłuszczowa |
50 |
|
Skóra sucha |
3030 |
|
Kości bez okostnej |
2x102 |
Opór elektryczny ciała ludzkiego dla prądów zmiennych niskiej częstotliwości
W przypadku, kiedy do tkanki przyłożymy zmienne napięcie U o ustalonej częstotliwości kątowej co, przez tkankę popłynie prąd zmienny o natężeniu I, takim że
![]()
gdzie Z oznacza opór pozorny tkanki (tzw. impedancję) i równa się
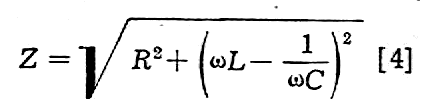
Jeżeli impedancja dotyczy tylko błony komórkowej (Zm), a wnętrze komórki jest wolne od impedancji, to można napisać
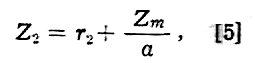
i
wtedy
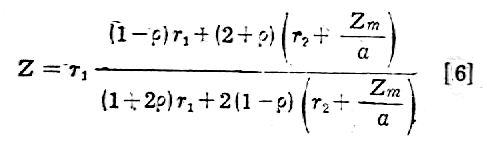
Jeżeli błona komórkowa ma pojemność Cm na jednostkę powierzchni i nie ma oporu upływności, to przy małych częstotliwościach otrzymamy: tylko opór rq:
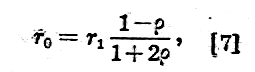
a przy dużych częstotliwościach
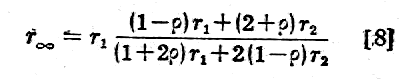
Pomiary r1 i r0 określają wielkość r, a pomiar rv określa wielkość r2, wewnętrzny opór właściwy komórki.
Przy pomiarach osiowego oporu elektrycznego pojedynczego włókna otrzymuje się dla bardzo dużych częstotliwości
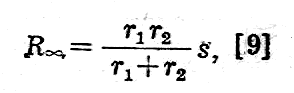
gdzie s oznacza odległość pomiędzy elektrodami.
Dla w=0 wyrażenie na opór (R0) składa się z dwóch składników - jednego podanego we wzorze [9] i drugiego bardziej skomplikowanego.
Doświadczalnie mierzy się Rv, R0 przy różnych odległościach s i stad wyznacza się r1, r2 i r4 i w ten sposób określa opory: wewnętrzny, zewnętrzny i błony komórkowej.
Dla częstotliwości pośrednich otrzymuje się na impedancje wzór:
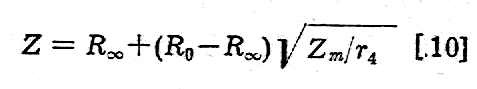
Stały prąd elektryczny, płynący pomiędzy elektrodami przyłożonymi do skóry zwierzęcia, napotyka w pierwszym rzędzie na opór elektryczny skóry. Opór reszty tkanek gra już wtedy rolę drugorzędna. W skórze suchej prąd płynie głównie przez kanaliki gruczołów potowych i niektórych tłuszczowych.
W skórze wilgotnej przewodnictwo elektryczne jest znacznie większe. Na skutek przesunięcia i lokalnych zmian koncentracji jonów występują w tkankach poddanych działaniu prądu stałego zjawiska polaryzacji elektrolitycznej, które zmniejszają wartość natężenia prądu.
Zjawisko to nie zachodzi dla prądów zmiennych i dlatego opór elektryczny tkanek dla prądu zmiennego jest znacznie mniejszy. Prąd stały używa się w lecznictwie do wprowadzania do organizmu różnego rodzaju jonów, do galwanizacji stabilnej i labilnej oraz kąpieli komorowych.
Aparat diatermiczny
Przenikanie prądów szybkozmiennych o dużym natężeniu, rzędu kilku amperów, przez organizm ludzki bez szkody dla niego zostało wykorzystane dla celów leczniczych. Mianowicie, jak wiemy, z przechodzeniem prądu elektrycznego związane jest wydzielanie się energii cieplnej w postaci ciepła Joule'a w ilości proporcjonalnej do kwadratu natężenia prądu. Przy prądach stałych i prądach zmiennych małej częstotliwości wydzielanie większej ilości energii cieplnej jest związane z przepływem prądu, który wywołuje w organizmie duże zmiany elektrochemiczne. Ponieważ prądy szybkozmienne nie powodują takich dużych zmian, można je stosować do ogrzewania narządów zewnętrznych i wewnętrznych ciała. Ta metoda leczenia nosi nazwę diatermii.
Diatermia pozwala na ogrzewanie poszczególnych części wewnętrznych organizmu, podczas gdy ogrzewanie ciepłem z zewnątrz może sięgać zaledwie na parę milimetrów pod skórę, głównie ze względu na szybkie rozprowadzenie ciepła przez obieg krwi.
Zabiegi diatermiczne wywołują przejściowy spadek ciśnienia krwi w tętnicach, rozszerzanie się naczyń krwionośnych itd. Stosuje się je w reumatyzmie, podagrze, zapaleniach stawów, chorobach ginekologicznych itd.
Zasadniczy schemat aparatu diatermicznego pokazany jest na ryc.3. Prąd zmienny z sieci o napięciu około 200 woltów wchodzi do pierwotnego uzwojenia transformatora T i wychodzi przetworzony na prąd 2000 woltów. Zasila on obwód oscylatora O o pojemności C1, iskierniku I i cewce S1, w którym to obwodzie powstają prądy wielkiej częstotliwości. Z obwodem oscylatora sprzężony jest indukcyjnie drugi obwód drgań, zawierający pojemność C2 cewkę S2, amperomierz cieplny A i elektrody, pomiędzy którymi znajduje się ciało pacjenta.
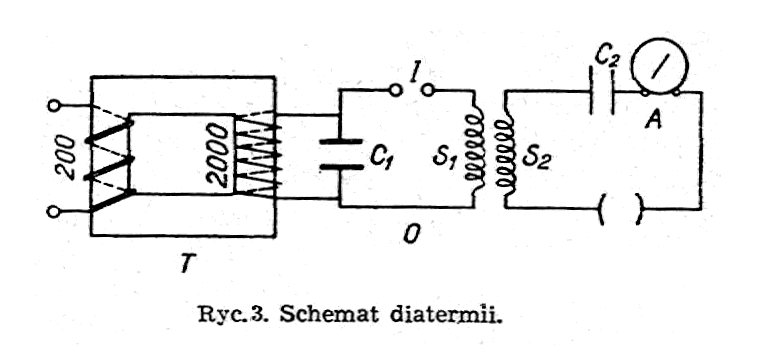
Rozróżniamy
obecnie dwa rodzaje prądów diatermicznych, zależnie od ich
częstotliwości drgań; diatermię długofalową (długość fali około 300
metrów, tzn. częstotliwość drgań około miliona razy na
sekundę) i diatermię krótkofalową (długość fali od 3 do 12
metrów, tzn. częstotliwość drgań od 25 do 100 milionów
razy na sekundę)
Obydwa rodzaje prądów diatermicznych działają bardzo różnie na poszczególne tkanki ciała Na przykład diatermia długofalowa ogrzewa przede wszystkim tkanki znajdujące się tuż pod skórą, później mięśnie, a najmniej kości; natomiast diatermia krótkofalowa ogrzewa tkanki bardziej równomiernie, ale najintensywniej kości i wątrobę.
Ryc.4 przedstawia schematycznie krzywe rozkładu temperatury w różnych tkankach w zależności od rodzaju diatermii Jak widzimy z ryciny, temperatury poszczególnych tkanek różnią się minimalnie w przypadku stosowania diatermii krótkofalowej (FK), a natomiast różnią się znacznie w przypadku stosowania diatermii długofalowej (FD), przy czym największa temperaturę osiąga tkanka tłuszczowa (Tł), mniejszą - tkanka kostna (K) i jeszcze mniejsze, ale już bliskie temperatury osiągają: skóra (S), wątroba (W), mózg (M) i mięśnie (Ms).
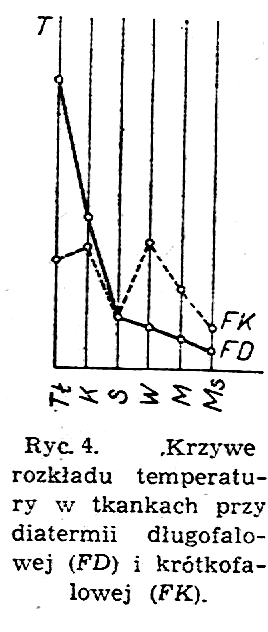
W
diatermii długofalowej przykłada się elektrody bezpośrednio do ciała
pacjenta, natomiast w diatermii krótkofalowej elektrody
znajdują się w pewnej odległości od ciała z tym, że musi się ono
znajdować pomiędzy nimi w polu elektrycznym kondensatora.
Fulguracja, zimna kaustyka, elektrowstrząsy
Prądy wielkiej częstotliwości mogą jednak wywołać, przy zbyt dużym wydzielaniu ciepła albo przy wyładowaniach iskrowych, szkodliwe spalanie tkanek. W nowszych czasach wyładowania iskrowe z transformatorów wielkiej częstotliwości zostały wykorzystane do odkażania świeżych płaszczyzn cięcia przy operacjach (tzw. fulguracja), szczególnie gdy chodzi o zniszczenie ewentualnych zawiązków raka. Wytwarzany przy tym ozon ma również pewną rolę odkażającą.
Poza tym stosuje się prądy wielkiej częstotliwości w elektrochirurgii przy przecinaniu tkanek (tzw. elektrotomia). W pewnym miejscu ciała pacjenta przykłada się wtedy elektrodę o dużej powierzchni, a w miejscu ciecia przykłada się drugą elektrodę w postaci wąskiego i cienkiego noża platynowego (lancetu). Elektrody włącza się do źródła prądu wielkiej częstotliwości i wtedy ze względu na bardzo dużą gęstość prądu (rzędu 104A/m2), jaka wytwarza się na ostrzu lancetu, tkanki zostają na bardzo wąskiej przestrzeni po prostu spalone; powstające przy tym iskierki są wielkości mikroskopijnej, tak że przy szybkim ruchu noża ślad cięcia jest minimalny. Jednocześnie następująca koagulacja przeciwdziała krwawieniu tkanek w miejscu ciecia. Taki lancet elektryczny nazywamy kauterem wielkiej częstotliwości albo zimnym kauterem.
Do najnowszych działów elektromedycyny, stosowanych praktycznie dopiero od kilkunastu lat, należą: elektrowstrząsy i elektronarkoza. W obydwu przypadkach zjawisko polega na oddziaływaniu prądu elektrycznego na tkankę mózgową pacjenta. Różnica działania jest natury ilościowej.
Przy elektrowstrząsach przepuszczamy prąd zmienny (albo jednokierunkowy modulowany) przez głowę pacjenta w ciągu bardzo krótkiego czasu (od 0,1 s do 4 s), przy tym czas jest regulowany dokładnie w każdym aparacie przez specjalny wyłącznik automatyczny. Napięcie takiego prądu jest najczęściej rzędu stukilkudziesięciu woltów (w niektórych aparatach dochodzi do 300 woltów), natężenie prądu może się wahać w granicach od 150 do 800 miliamperów.
Przy elektronarkozie stosuje się napięcia mniejsze, tak żeby prąd elektryczny miał natężenie rzędu kilkudziesięciu do stu miliamperów, przy tym często można stosować również obok prądu zmiennego i prąd stały. Czas zabiegu jest przy tym dłuższy (rzędu kilku minut).
Aparaty do elektrowstrząsów i do elektronarkozy są zaopatrzone w przyrządy pomiarowe (amperomierze, woltomierze i omomierze) w urządzenia zabezpieczające pacjenta i obsługę przed zwiększeniem napięcia, przed oparzeniami i porażeniami w razie nagłego oderwania się elektrod od skóry pacjenta w czasie zabiegu.
Działanie mikrofal (fal radarowych i telewizyjnych)
Wraz z rozwojem radiotechniki, radiolokacji i teletechniki oraz licznymi zastosowaniami tych dziedzin w technice, w przemyśle i w wojskowości powstało bardzo poważne zagadnienie szkodliwego oddziaływania prądów wysokiej częstotliwości na żywy organizm. Szczególnie jest to o tyle ważne, że zaczęto wytwarzać źródła tych prądów o bardzo dużej mocy dochodzącej do 100 megawatów i o częstotliwości rzędu dziesiątków tysięcy megaherców. W związku z tym liczba ludzi znajdujących się w zasięgu oddziaływania takiego promieniowania elektromagnetycznego w. cz. i narażonych na jego oddziaływanie szybko wzrasta.
Obecnie ustalane są już normy dopuszczalnego maksymalnego dawkowania człowieka takim promieniowaniem w zależności od częstotliwości promieniowania od długości jednorazowego napromieniowania itd.
W przemyśle, w technice i w medycynie stosuje się promieniowanie w. cz. w granicach częstotliwości od 3 MHz (długość fali 100 m) do 3x105 MHz (długość fali 1 mm).
Pola elektromagnetyczne w.cz. w granicach od około 3 do 30 MHz stosuje się w przemyśle i w technice do termicznej obróbki metali i dielektryków, do suszenia, klejenia, spawania, polimeryzacji, sterylizacji żywności, niszczenia szkodników, do łączności radiowej itd.
Pola elektromagnetyczne o częstotliwości w granicach od 30 do 300 MHz stosuje się w łączności radiowej i telewizyjnej.
Specjalnie dużo uwagi poświęcono tzw. mikrofalom, których zakres długości fali wynosi 10-1 - 100 cm, a częstotliwość 3x102- 3x105 MHz.
Do tego zakresu należą fale radarowe i fale telewizyjne. Mikrofale graniczą z jednej strony z falami radiowymi ultrakrótkimi (o częstotliwości od 30 do 300 MHz), z drugiej strony z promieniowaniem podczerwonym (o częstotliwości od 3x106 do 4x108 MHz).
Cały zakres opisanego wyżej pola elektromagnetycznego w. cz. znajduje również zastosowanie w medycynie, między innymi w diatermii. Poza tym stanowi duży problem dla bezpieczeństwa i higieny pracy (bhp). Poza bezpośrednim działaniem fizjologicznym tych pól należy się jeszcze liczyć z działaniem promieniowania rentgenowskiego, jakie może być wytworzone ubocznie w generatorach mikrofalowych.
ELEKTRYCZNE WŁAŚCIWOŚCI KOMÓRKI
Właściwości błony komórkowej.
Komórki żywego organizmu są źródłem siły elektromotorycznej (SEM). Siła ta powstaje na skutek wędrówki jonów i zmiany ich koncentracji w różnych miejscach komórki. Takie przesunięcia jonów wywołują powstanie w tych miejscach różnicy potencjałów elektrycznych, która w pewnym momencie może wywołać impuls prądu elektrycznego przesuwający się wzdłuż tkanki i powodujący wyrównanie tych potencjałów
Z tego względu rozróżniamy w fizjologii potencjały spoczynkowe i tzw. potencjały czynnościowe towarzyszące impulsom prądu. Różnica potencjałów spoczynkowych powstaje najczęściej na skutek różnicy ruchliwości jonów przy ich przechodzeniu przez półprzepuszczalne błony komórek
Z tego względu obserwuje się na ogół różnicę potencjałów pomiędzy wewnętrzną częścią komórki i jej otoczeniem, a wiec po dwóch stronach błony komórkowej. Błona taka stanowi zatem pewnego rodzaju kondensator elektryczny z warstwa dielektryka w środku, który może ulec naładowaniu do różnicy potencjałów rzędu od ułamka do kilkuset miliwoltów.
Jest to napięcie bardzo małe, ale ze względu na znikomo małą grubość błony komórkowej (rzędu od kilkudziesięciu do kilku tysięcy angstremów) mogą w niej powstawać pola elektryczne o natężeniu setek tysięcy wotów na centymetr.
Opór elektryczny błony komórkowej jest rzędu 107 W/m2 lub 1,3x1012 W/m3, jej pojemność elektryczna wynosi około 1,8x104 mF/m2. Opór elektryczny plazmy i płynów międzykomórkowych wynosi około 5x107 W/m3.
Taki stan błony nazywamy stanem spolaryzowanym. Jeżeli następuje rozładowanie tego kondensatora, mówimy, że nastąpiło pobudzenie komórki, a następnie jej depolaryzacja, tzn. wyrównanie potencjałów.
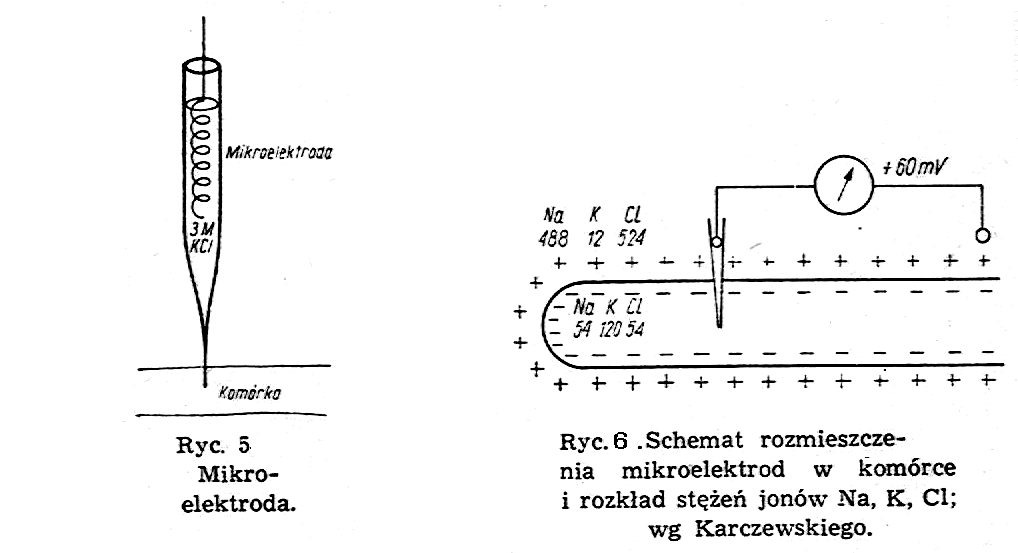
Badania doświadczalne potencjałów czynnościowych przeprowadza się za pomocą mikroelektrod. Mikroelektroda jest to rurka zakończona cienką kapilarą o średnicy rzędu jednego mikrometra (ryc.5), wypełniona 3 M KCl. W rurze znajduje się zwinięty spiralnie drucik srebrny pokryty AgCl2.
Mikroelektrodę wkłuwa się za pomocą mikromanipulatora do wnętrza komórki, a drugą elektrodę, uziemioną umieszcza się na zewnątrz komórki w płynie fizjologicznym (ryć. 6).
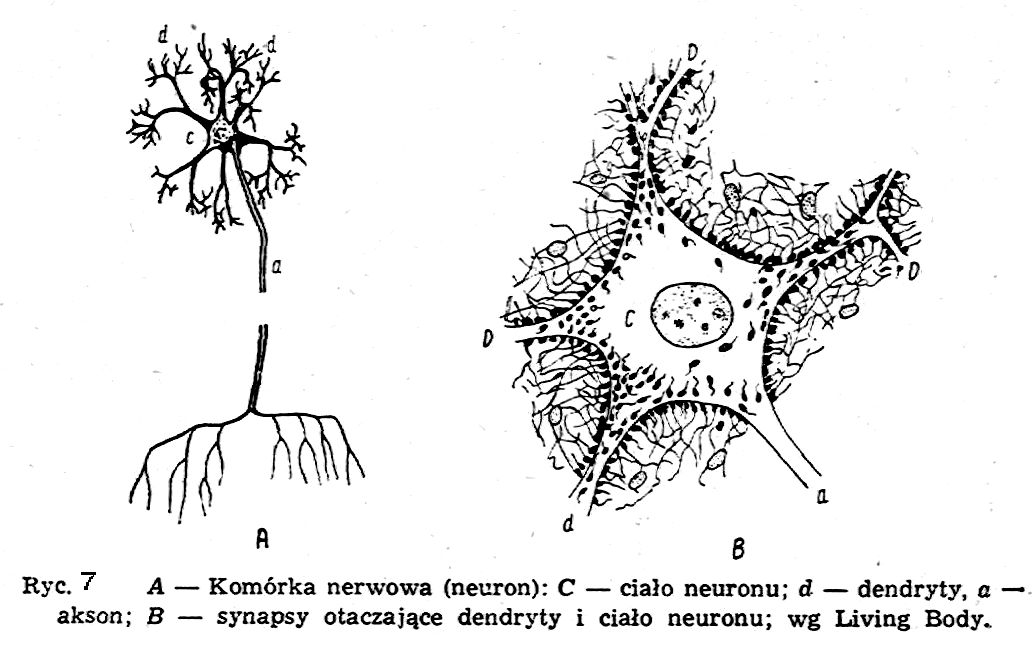
Mikroelektrodę dołącza się do jakiegoś czułego miliwoltomierza, elektrometru czy oscyloskopu elektronowego. Dla zobrazowania procesów fizycznych zachodzących w żywej komórce musimy najpierw przypomnieć najważniejsze cechy fizyczne żywej komórki. Jako przykład weźmiemy komórkę nerwową, tzw. neuron; jest to jedna z najważniejszych komórek w organizmie; dokonuje ona nie tylko przemiany materii, ale również wytwarza impulsy elektryczne, transformuje je, przewodzi i dostarcza do innych komórek nerwowych i mięśniowych.
Neuron.
Neuron składa się z niewielkiej części centralnej (tzw. ciała neuronu) (ryc.7) i z wielu wypustek, tzw. d e n d r y t ó w, wychodzących z tej części na zewnątrz i rozgałęziających się; poza tym istotną częścią neuronu jest tzw. a k s o n, długa pojedyncza wypustka, zakończona również odgałęzieniami. Akson odgrywa rolę kabla elektrycznego, tzn. przewodnika elektrycznego owiniętego warstwami izolatora. Średnica aksona jest rzędu 1-20 mikronów, podczas gdy jego długość może dochodzić do rzędu metrów.
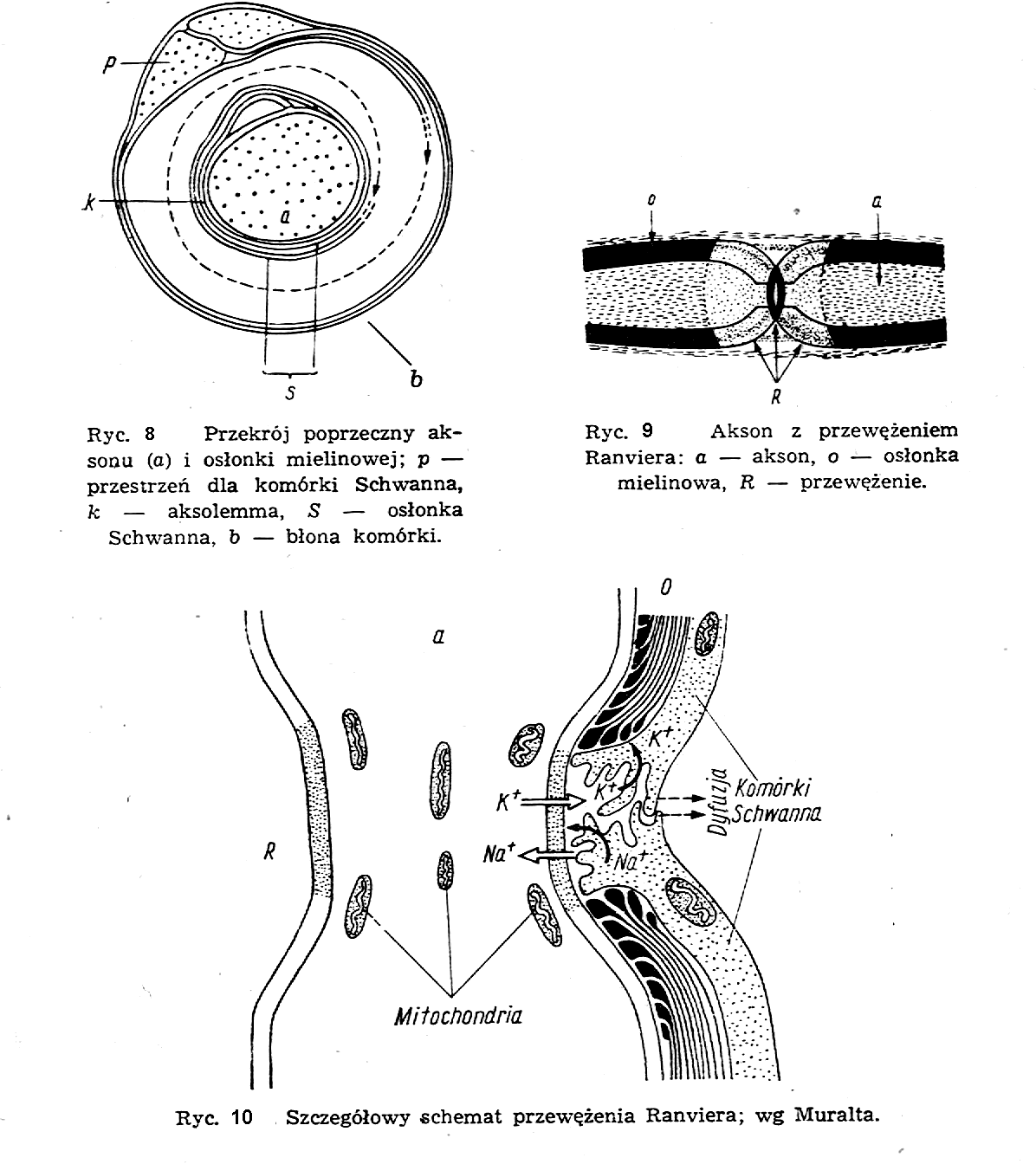
Błona komórkowa otaczająca akson składa się z kilku warstw. Warstwa wewnętrzna, najbliższa środka, tzw. błona pobudliwa, jest zbudowana w postaci kondensatora; jego okładki są utworzone z dwóch warstw białka, a dielektrykiem jest warstewka tłuszczu, znajdująca się pomiędzy nimi. Grubość błony pobudliwej jest rzędu kilkudziesięciu angstremów. Na zewnątrz od tej błony znajdują się inne warstwy osłaniające akson (ryc. 8): osłonka mielinowa, osłonka nerwowa, osłonka Schwanna i tzw. endoneurium.
Osłonka mielinowa składa się z krótkich odcinków włókna zbudowanego z białek i z fosfolipidów o długości od 0,5 do 3 mm, owijających akson. Zwoje osłonki mielinowej nie otaczają szczelnie aksonu na całej jego długości, ale tworzą pewne przerwy, które noszą nazwę przewężeń Ranviera (ryc 9); w tych przerwach akson jest odsłonięty. Na ryc. 10 pokazany jest szczegółowy schemat przewężenia Ranviera.
Na końcu aksonu znajdują się rozgałęzienia również nie osłonięte osłonką mielinowa i zakończone zgrubieniami, wewnątrz których znajdują się cząstki acetylocholiny o średnicy rzędu kilkuset angstremów.
Rozkład potencjałów w komórce nerwowej
Badania wielkości i rozkładu potencjałów w komórkach wykazują, że na ogół wnętrze komórki ma potencjał ujemny w stosunku do potencjału płynu pozakomórkowego, przy tym wartość tego potencjału w komórkach nerwowych jest rzędu 90 mV, w innych komórkach jest przeważnie mniejsza.
Poza
tym wykazały one, że przyczyną powstawania tej różnicy
potencjałów jest głównie różnica w stężeniach
jonów sodu, potasu i chloru po obu stronach błony komórkowej.
Mianowicie, wewnątrz komórki stężenie jonów sodu (Na+)
jest około 10 razy niższe niż na zewnątrz, stężenie jonów
chloru (Cl-) jest od 10 do 65 razy niższe, a stężenie jonów
potasu (K+) jest około 10 do 50 razy wyższe niż na zewnątrz.
Wartości te wahają się w dosyć dużych granicach i są uzależnione od przepuszczalności błony komórkowej dla różnych jonów, przy tym półprzepuszczalność błony może być uwarunkowana różnymi czynnikami, np. duża ilość białka w komórce wpływa na powiększenie różnicy w koncentracji jonów.
Poza tym błony komórkowe różnych komórek wykazują niekiedy bardzo selektywne właściwości przepuszczania różnych rodzajów jonów, np. powłoki czerwonych krwinek (erytrocytów) przepuszczają głównie aniony, membrany komórek nerwowych przepuszczają głównie potas, a nie przepuszczają sodu itd.
W tabeli poniżej podany jest przykład stężenia jonów w aksoplaźmie włókna Loligo i w płynie pozakomórkowym (w materiale świeżo spreparowanym).
Stężenie jonów K+, Na+ i Cl- [w mol/kg] wewnątrz (a) i na zewnątrz (b) komórki
|
|
a |
b |
Stosunek stężeń |
|---|---|---|---|
|
K+ |
410 |
10 |
41 |
|
Na+ |
49 |
460 |
0 |
|
Cl- |
40 |
540 |
0.074 |
Różnica w stężeniach jonów po obydwu stronach błony komórkowej powoduje powstawanie w tych miejscach różnicy potencjałów elektrycznych oraz przeciwdziałające im działanie siły dyfuzji, która dąży do wyrównania stężeń. Różnica potencjałów, która równoważy siłę dyfuzji w stanie równowagi dynamicznej przy danej różnicy stężeń nosi w fizjologii nazwę potencjału równowagi dla danego jonu Potencjał równowagi (E) określa się z równania Nernsta:
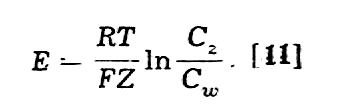
gdzie
T oznacza temperaturę w skali bezwzględnej, F - stałą Faradaya, Z -
wartościowość danego jonu, Cz i Cw - stężenia
jonów danego rodzaju na zewnątrz i wewnątrz komórki.
Dla jonów jednowartościowych (Z = 1) w temperaturze ciała ludzkiego (t = 37°C, T = 310°K) wzór [11] przybiera formę prostszą
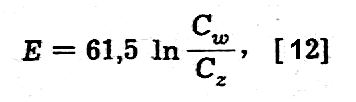
co
np. dla potasu Cw/Cz= 41 daje wartość 95 mV
bardzo bliską wartości doświadczalnej.
Opisany wyżej prosty mechanizm wytwarzania potencjałów elektrycznych w komórce nasuwa jednak jeszcze wiele niejasności i wątpliwości. Z tego względu rozpatrywano również możliwości szeregu innych procesów, które mogą tłumaczyć zjawiska elektryczne obserwowane w żywym organizmie. Pod uwagę brano tutaj niektóre procesy znane z innych działów fizyki, np. wpływ przenoszenia swobodnych elektronów, swobodnych protonów, mechanizm przenoszenia nośników prądu w półprzewodnikach elektronowych czystych i domieszkowych, właściwości elektryczne warstw podwójnych itd.
Na przykład Davies zwrócił uwagę, że, jak wiadomo, membrana komórkowa ma charakter lipoproteinowy, przy czym warstwa lipidów styka się z wodnymi roztworami wewnątrz i na zewnątrz komórki. Dlatego przy rozpatrywaniu elektrycznych potencjałów komórkowych biofizyk powinien znać właściwości takich powierzchni rozdziału.
Energia jonów na granicy rozdziału dwóch faz zależy od położenia (orientacji) dipoli w obu fazach. W wodzie energia jonów jest stosunkowo duża, w lipidach mała. Między fazami lipidową i wodna nie ma ostrego przejścia, istnieje strefa przejściowa. Dla przejścia drobiny z fazy wodnej w lipidową trzeba pokonać energię hydratacji. Przy przejściu z fazy lipidowej w wodna energia solwatacji jest dużo mniejsza (energia przejścia odpowiada długości odcinka). Ta różnica energii prowadzi do powstawania gradientów koncentracji i określa wielkość współczynnika rozdziału (rozkładu).
Teoretycznie różnica koncentracji, pojawiająca się w następstwie przenoszenia energii jonów poprzez membranę komórkowa, może prowadzić do powstawania potencjałów elektrycznych znacznych wielkości. Taka energetyczna bariera uwarunkowuje małą przenikliwość, duży elektryczny opór i mniejszą dyfuzję przez membranę. Wszystkie te właściwości dadzą się obserwować w membranach żywych komórek.
Istniała również teoria oparta na teorii kwasowo-zasadowych potencjałów, w której wykazano, że dużo bioelektrycznych zjawisk można wyjaśnić przenoszeniem protonów. Wymiary protonów są znacznie mniejsze od rozmiarów jonów w metalu, natomiast gęstość ładunków protonów jest o wiele większa od gęstości ładunku tych jonów. Proton może przenosić się od drobiny do drobiny, podobnie jak elektron. Przewodnictwo takiego typu obserwuje się w kwasie siarkowym, a także w kwasach i w innych substancjach znajdujących się w stanie stałym.
Przy koncentracjach reagujących substancji obserwowanych w żywych komórkach (tkankach), różnica potencjałów może być równa kilkaset mV. Można więc powiedzieć, że powstawanie potencjałów bioelektrycznych może być uwarunkowane procesami protonowo-chemicznymi Istnieje jeszcze kilka innych teorii powstawania potencjałów bioelektrycznych, ale nie będziemy ich tu omawiali.
Przenoszenie bodźców elektrycznych w komórkach.
Bodźcami pobudzającymi komórkę mogą być procesy chemiczne, mechaniczne, elektryczne, świetlne lub cieplne. W badaniach doświadczalnych w biofizyce i w fizjologii stosuje się najczęściej bodźce elektryczne, wprowadzając do wnętrza komórki mikroelektrodę i doprowadzając do niej odpowiedni potencjał. Jeżeli ten potencjał jest ujemny, tzn. jeżeli mikroelektroda jest katodą, to przy pewnej wartości tego potencjału (tzw. wartości progowej) może nastąpić depolaryzacja błony komórkowej i występuje w komórce gwałtowny wzrost potencjału (tzw. potencjał czynnościowy) w postaci ostrego wierzchołka (ryc. 11) na krzywej rozkładu potencjału.
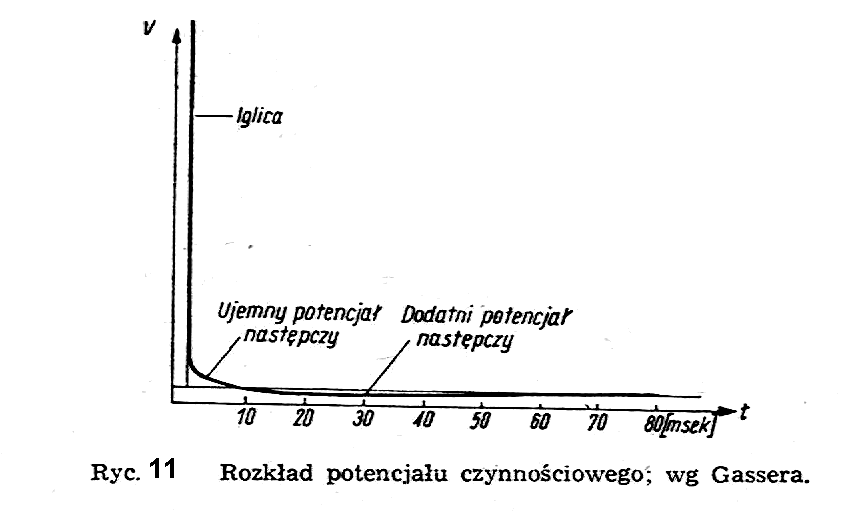
Odpowiada to jak gdyby "przebiciu elektrycznemu" błony komórkowej, gdyż jej opór elektryczny zmniejsza się w danym miejscu z wartości około 107 W/m2 do wartości około 2,5x 105 W/m2.
W miejscu "przebicia" przepuszczalność błony komórkowej dla jonów sodu staje się prawie całkowita, dzięki czemu następuje wyrównanie stężenia jonów sodu po obydwu stronach błony komórkowej, tzn. że jony sodu z zewnątrz "wlewają" się do wnętrza komórki niosąc ze sobą dodatni ładunek elektryczny, który znosi ujemny potencjał istniejący uprzednio wewnątrz komórki. W chwilę po tym rozpoczyna się inny proces, mianowicie gwałtowna dyfuzja jonów potasu z wnętrza komórki na zewnątrz, dzięki czemu zaczyna częściowo wzrastać różnica potencjałów pomiędzy wnętrzem komórki i przestrzenią pozakomórkową. Obydwa procesy zachodzą zgodnie z działaniem sił gradientu elektrochemicznego, tzn. powodują rozładowanie ogniwa stężeniowego w komórce. Na ryc.12 pokazane są zmiany w czasie przepuszczalności błony komórkowej dla jonów sodu p(Na+) i dla jonów potasu p(K+) i wytworzony w ten sposób potencjał czynnościowy (Vc).
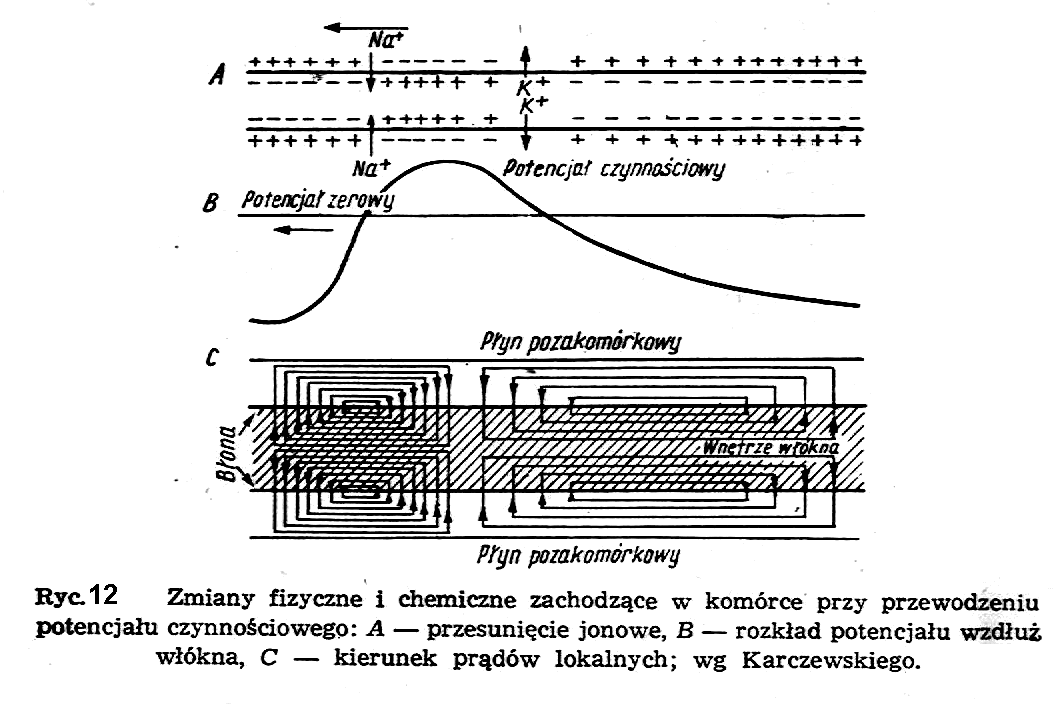
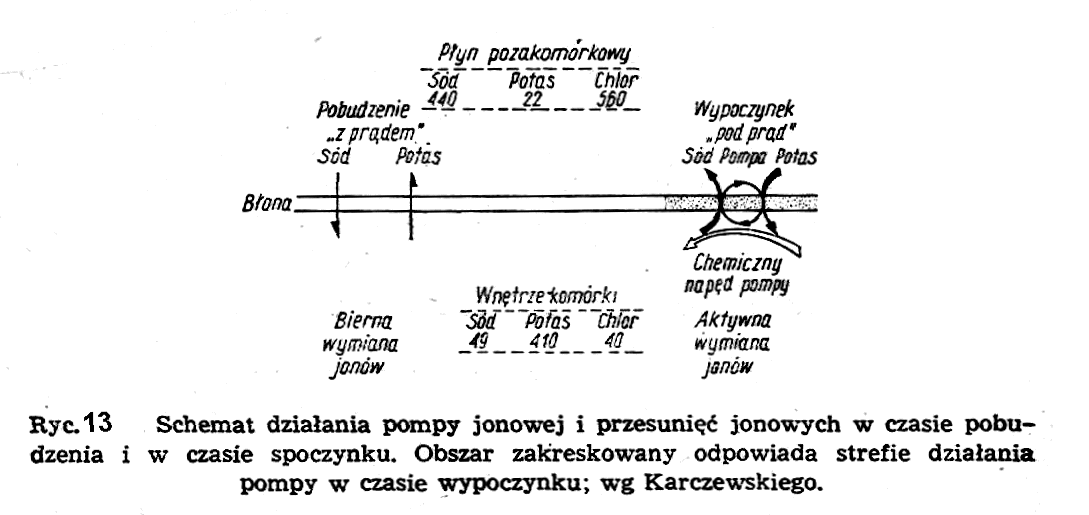
Cały proces zachodzi w czasie rzędu paru milisekund. Ażeby przywrócić w komórce poprzedni stan typu ogniwa stężeniowego, należy sobie wyobrazić jakiś mechanizm, który przepycha jony sodu z powrotem na zewnątrz komórki, a jony potasu do jej wnętrza w obydwu przypadkach wbrew działaniom gradientów elektrochemicznych, tzn. powiększa różnice stężeń jonów każdego rodzaju po obydwu stronach błony komórkowej. Taki mechanizm nosi nazwę ogólnie pompy jonowej, a w szczególności pompy sodowej lub pompy potasowej (ryc. 13).
Jest on umieszczony całkowicie w błonie komórkowej, ale czerpie energię z protoplazmy. Energia ta powstaje z degradacji wysokoenergetycznych związków chemicznych w obecności tlenu, który jest konieczny dla działania tego mechanizmu.
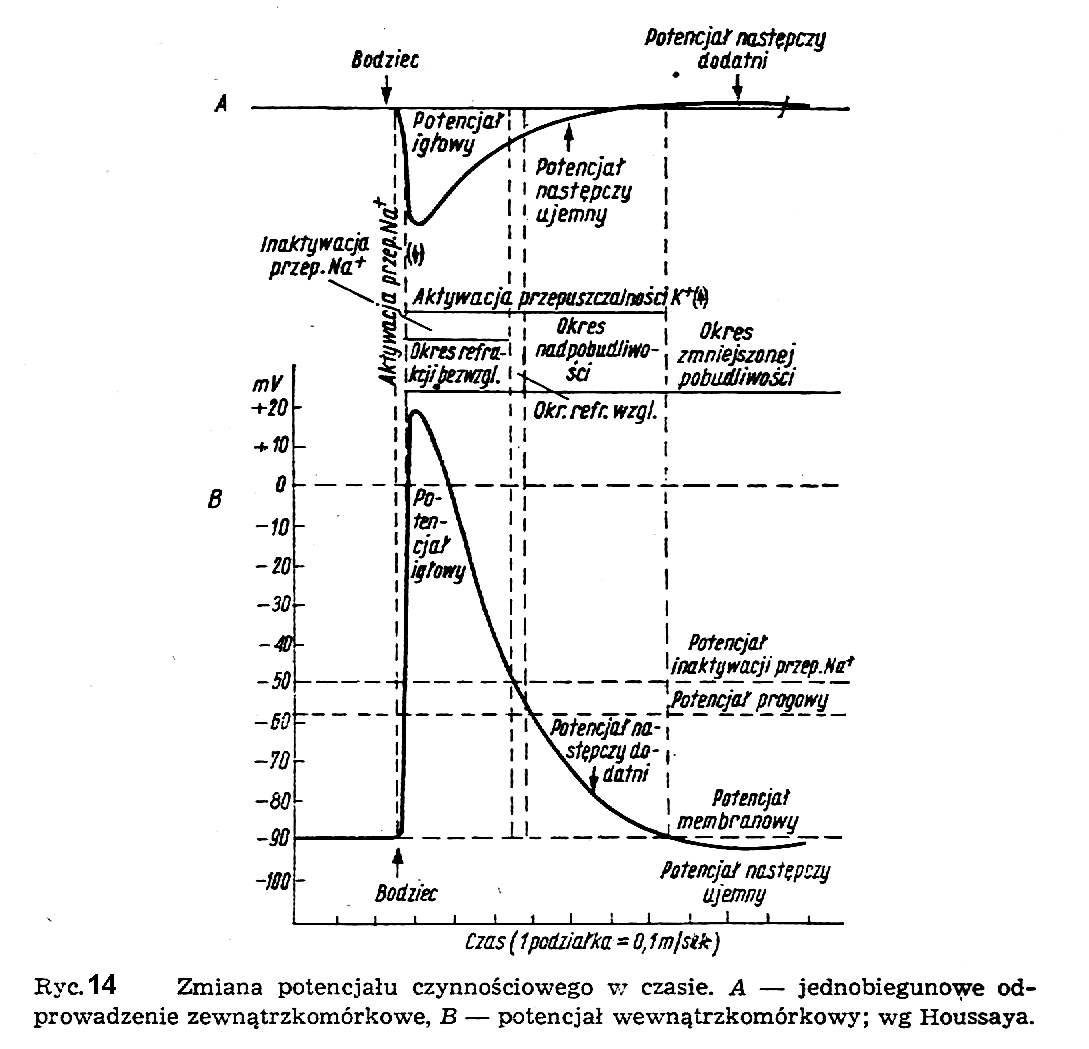
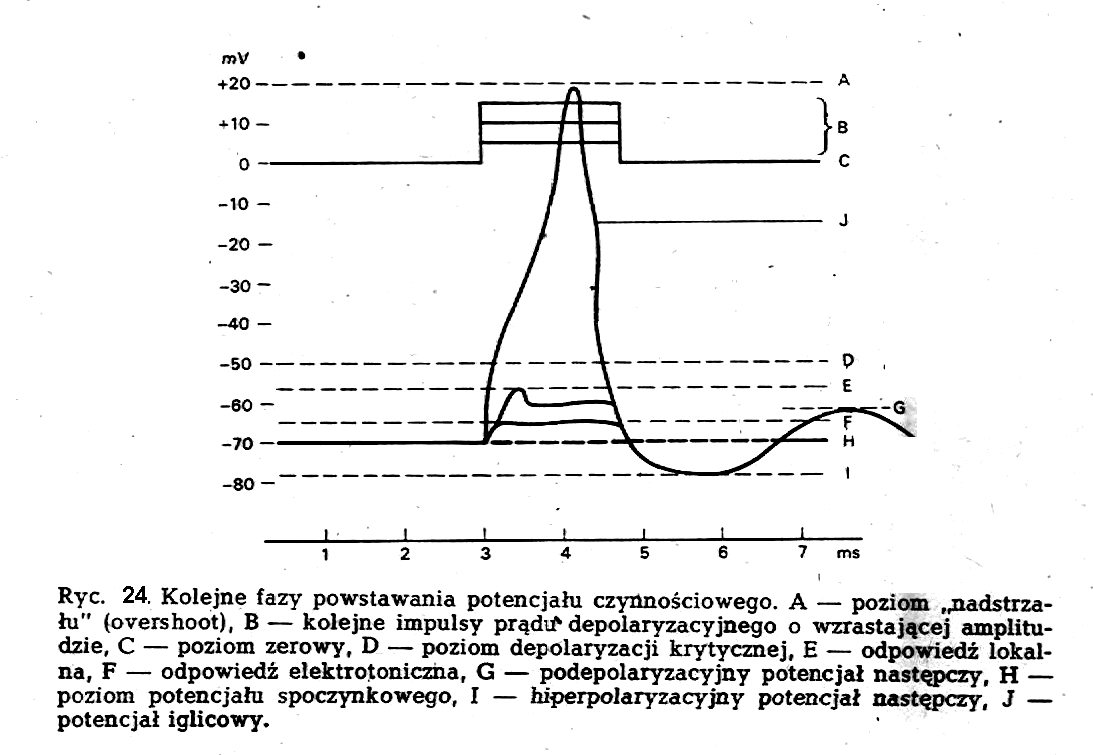
Dalsze badania nad przebiegiem potencjału czynnościowego w czasie wykazały, że jest on funkcją bodźca i ma kształt pokazany na ryc. 14.
Po pierwszym, gwałtownym wzroście (w postaci iglicy) przy depolaryzacji błony komórkowej, następuje powolny spadek aż do wartości poniżej wartości początkowej i powrót do stanu wyjściowego. Poszczególne odcinki krzywej potencjału czynnościowego noszą nazwę potencjału igłowego i potencjałów następczych: ujemnego i dodatniego.
Jak widać z powyższego opisu procesu pobudzania, mechanizm potencjału" czynnościowego jest bardzo skomplikowany, a trzeba jeszcze podkreślić, że w dużej mierze jeszcze hipotetyczny i niedostatecznie ugruntowany doświadczalnie pomimo bardzo wielu prac. prowadzonych przez wielu autorów różnymi metodami współczesnej fizyki, chemii i fizjologii. Część prac nad mechanizmem wędrówki jonów przez błonę komórkową wykonano za pomocą metod radioizotopowych stosując promieniotwórcze izotopy sodu i potasu. Powstanie potencjału czynnościowego jest tylko pierwszą częścią procesu przenoszenia impulsów pobudzenia przez komórki. Zmiany potencjału wewnątrz komórki są źródłem siły elektromotorycznej prądu elektrycznego, który może przepływać przez nerwy i włókna mięśni pomiędzy miejscami o różnym potencjale. W zależności od rodzaju przewodnika przewodzenie może być ciągłe albo przerywane (skokowe).
Przewodzenie ciągłe powstaje w bezrdzennych włóknach, nerwowych, we włóknach mięśni szkieletowych i gładkich oraz w komórkach mięśnia sercowego. Przewodzenie skokowe odbywa się we włóknach mielinowych, w których występują przewężenia Ranviera. Szybkość przewodzenia skokowego jest większa niż szybkość przewodzenia ciągłego. Proces wytworzenia potencjału czynnościowego pod wpływem bodźca zewnętrznego i przekazania go dalej wzdłuż nerwu czy włókna mięśniowego zależy nie tylko od wielkości (siły) tego bodźca, ale i od czasu jego trwania i od poprzedniego stanu komórki.
W fizjologii przyjmuje się na ogół prawo "wszystko albo nic", tzn. prawo, które mówi, że dany bodziec działa w sposób warunkowy. Jeżeli jest on dostatecznie duży w stosunku do progu pobudliwości komórki, to wywoła impuls, jeśli niniejszy od tego progu, to nie wywoła nic. Istnieją jednak przypadki, że bodźce o mniejszej wartości wywołują pewien stan podprogowy w komórce, który zmienia jej zachowanie w stosunku do następnego takiego bodźca. Drugim parametrem grającym rolę przy pobudzaniu komórki jest czas działania bodźca; im krótszy czas działania, tym bodziec musi być silniejszy. Wreszcie trzecim parametrem jest uprzedni stan komórki. Komórka wyładowana (depolaryzowana) nie reaguje przez pewien czas na bodźce, a zatem wykazuje jak gdyby "pewien czas martwy". Jest on rzędu 0,2 do 2 milisekund. Taki stan komórki nosi w fizjologii nazwę refrakcji bezwzględnej. W przypadku, kiedy bodźce przychodzą za często, komórka może zareagować tylko częściowo albo też wymaga zwiększonej siły bodźców dla normalnego zareagowania. Taki sposób reagowania nosi nazwę refrakcji względnej. Mechanizm przewodzenia bodźców we włóknie bezrdzeniowym jest bardzo skomplikowany. Nie znajduje on wyraźnej analogii w znanych w fizyce mechanizmach przewodzenia prądów elektrycznych w metalach, w półprzewodnikach czy w elektrolitach. Niekiedy porównuje go się raczej do mechanizmu działania lontu, w którym, jak wiadomo, płomień przenoszony jest powoli z miejsca do miejsca, czerpiąc energię z nagromadzonego w danym miejscu materiału palnego.
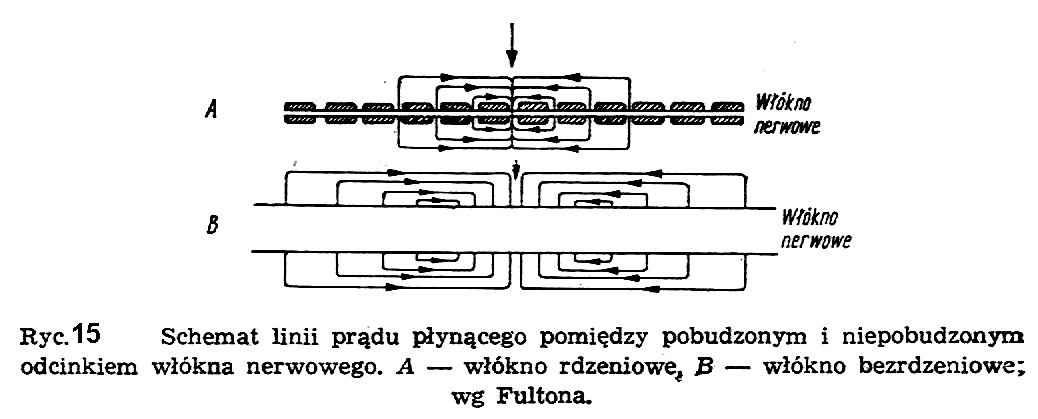
Na ryc. 15 pokazany jest schematycznie obraz hipotetycznego mechanizmu przewodzenia bodźca wzdłuż włókna rdzeniowego (A) i bezrdzeniowego (B). Po lewej stronie rysunku jest stan normalny włókna, w części środkowej następuje pobudzenie (zmiana potencjału czynnościowego), w części prawej następuje repolaryzacja i powrót do stanu normalnego.
Zatem przed każdym potencjałem czynnościowym wędruje we włóknie fala pobudzenia, która obniża próg pobudliwości następnej warstwy włókna. Tego rodzaju prędkość przewodzenia bodźców jest rzędu 0,5 do 2 m/s. Włókna bezrdzeniowe znajdują się w układzie wegetatywnym.
Inny mechanizm przewodzenia bodźców elektrycznych zachodzi we włóknach rdzeniowych. Jest to mechanizm typu skokowego. Jak wiemy przewodzenie w komórce nerwowej odbywa się głównie wzdłuż aksonu, który sam jest dobrym przewodnikiem elektryczności (jego opór właściwy wynosi około 1 Wm), a jest osłonięty na przeważającej swojej części przez osłonę nielinową, która jest bardzo dobrym izolatorem (jej opór właściwy jest rzędu 8x107 Wm. Osłonka mielinowa ma szereg przerw, w których akson jest prawie odsłonięty i ulega przewężeniu. Mechanizm przewodzenia bodźców wewnątrz osłoniętych części aksonu pomiędzy przewężeniami Ranviera jest podobny do opisanego wyżej mechanizmu przewodzenia w układach bezrdzeniowych, natomiast w przewężeniach Ranviera obwód prądu zamyka się ponad osłonką nielinową wytwarzając nowy potencjał czynnościowy w następnym przewężeniu. A więc potencjał czynnościowy jak gdyby przeskakuje z jednego przewężenia Ranviera do drugiego, przy tym szybkość takiego przenoszenia bodźców jest zależna od rodzaju włókien nerwowych.
Rozróżnia się trzy rodzaje włókien nerwowych o następujących właściwościach elektrycznych: grupę A z podgrupami a, (a, b,g o średnicy rzędu 1-20m , prędkości przewodzenia rzędu 100-150 m/s i o czasie martwym rzędu 0,5 m; grupę B o średnicy około 3 m, o prędkości przewodzenia około 10 m i o czasie martwym rzędu 1,2 m; grupę C - najcieńsze, bezrdzeniowe, o prędkości przewodzenia około 1 m i o czasie martwym rzędu 2 m s.
Nerw jest złożony z różnych grup wielu tysięcy poszczególnych włókien nerwowych, tak że bodziec elektryczny wędruje różnymi drogami, dając w sumie w miejscu doprowadzenia złożony potencjał czynnościowy nerwu. Przekazywanie bodźców elektrycznych z jednego ośrodka do drugiego odbywa się najczęściej za pomocą dwóch lub więcej neuronów; zatem droga przewodzenia bodźca składa się z następujących elementów: dendryt, ciało neuronu, akson, zakończenie aksonu, tzw. synapsa, dendryt drugiego neuronu itd.
Synapsa jest elementem nie zupełnie jeszcze poznanym. Znajduje się ona pomiędzy dwoma neuronami. Zakończenie jednego neuronu od początku drugiego neuronu dzieli przerwa o szerokości około 700 A. W przerwie tej znajdują się czynne substancje chemiczne, takie jak acetylocholina, adrenalina i serotonina. Noszą one nazwę mediatorów. Poszczególne drobiny mediatora zostają uwolnione z tzw. pęcherzyków synaptycznych przez energię potencjału czynnościowego, przedostają się przez przerwę synaptyczną i działają na błonę osłaniającą drugi neuron. Działanie to wywołuje szereg drobnych miniaturowych potencjałów postsynaptycznych, które w sumie depolaryzują błonę i wytwarzają w niej potencjał czynnościowy.
Proces ten jest o tyle bardziej skomplikowany, że do błony jednego neuronu mogą dochodzić jednocześnie potencjały czynnościowe z tysięcy innych neuronów, przy tym niektóre z tych potencjałów mogą wywołać działanie przeciwne, hamujące pobudzenie. Synapsa wykonuje zatem szereg bardzo skomplikowanych czynności: segregujących, wybiórczych i sumujących bodźce i przekazujących je dalej, do następnego neuronu czy ostatecznie, do mięśnia.
W tym ostatnim przypadku bodziec trafia nie do kolejnej synapsy, ale do płytki końcowej, w której odbywają się procesy podobne jak w synapsie z udziałem odpowiednich mediatorów, tylko efekt końcowy jest inny, mianowicie skurcz mięśnia, a więc wykonanie pracy mechanicznej.
Każdy pojedynczy neuron ruchowy może pobudzić całą grupkę włókien mięśniowych, tzw. jednostkę ruchową. Wartość potencjału czynnościowego jednostki ruchowej jest stosunkowo mała, wynosi zaledwie około 0,5 m, czas trwania tego potencjału wynosi około 12 milisekund. Droga przewodzenia bodźców elektrycznych przez układ neuron, synapsę, mięsień jest jednokierunkowa, tzn. że w drugą stronę bodźce w tym układzie nie przechodzą, głównie z uwagi na działanie synapsy. Ponieważ jednak pobudzenie mięśnia jest z reguły powiązane z wysłaniem bodźca czuciowego w drogę powrotną do ośrodkowego układu nerwowego, musi istnieć również osobna linia przewodzenia tego bodźca dla zasygnalizowania wykonania pierwotnego polecenia, ewentualnego skorygowania go czy wreszcie wykazania uczucia bólu.
W tym powrotnym układzie przewodzącym główna rolę na początku odgrywają tzw. receptory, specjalne komórki, czy zakończenia nerwowe wyczulone selektywnie na poszczególne rodzaje bodźców: mechanicznych, akustycznych, chemicznych, świetlnych, elektrycznych itd. (ryc.17).
Specjalną grupę/receptorów stanowią komórki przestrzegające przed uszkodzeniem czy zniszczeniem organizmu i sygnalizujące takie objawy przez wywoływanie bólu.
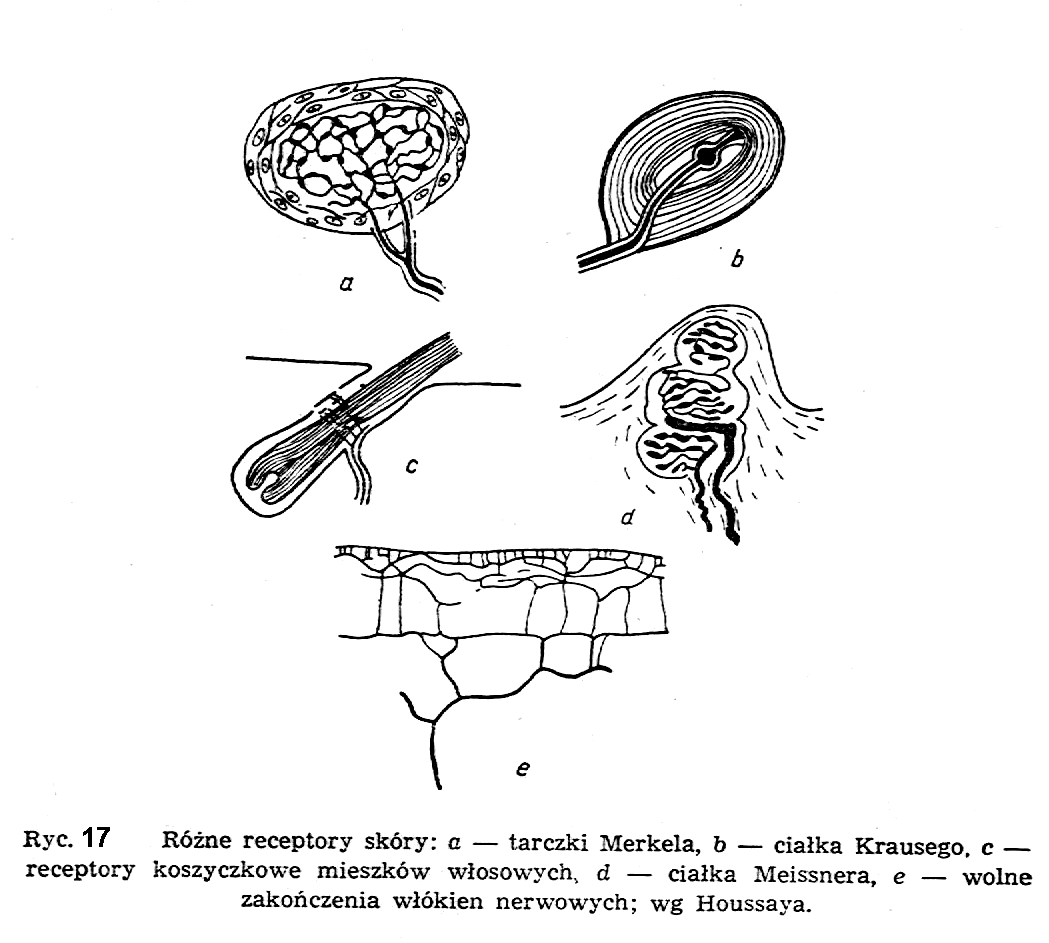
Istnieje bardzo wiele różnych rodzajów receptorów o różnej strukturze i różnych zadaniach funkcjonalnych (tzw. mechanoreceptory, chemoreceptory, termoreceptory itd.). W pobudzonym receptorze pojawia się tzw. potencjał generujący, który wysyła bodźce elektryczne pojedyncze albo grupowe w zależności od rodzaju i wielkości pobudzenia. Częstotliwość wysyłania tych bodźców elektrycznych nerwu charakteryzuje rodzaj przesyłanej informacji.
Można zatem powiedzieć, że informacje pochodzące z receptorów są kodowane za pomocą modulacji częstotliwości. Jak wiemy z innych dziedzin nauki, taki sposób przekazywania informacji jest znacznie dokładniejszy niż przekazywanie informacji za pomocą modulacji amplitudy i podlega znacznie mniejszym zniekształceniom. Reasumując rozważania ostatnich paragrafów, można powiedzieć, że droga, jaką przebiegają potencjały czynnościowe w ustroju, rozpoczyna się od źródła pierwszej podniety, które znajduje się najczęściej w receptorze obwodowym, przebiega zawiłą drogę poprzez dendryty, ciało neuronu, akson, synapsę, inne neurony, mięsień i dochodzi poprzez receptory i neurony do mózgu. Cały ten proces trwa zaledwie ułamek sekundy.
BUDOWA KOMÓRKI NERWOWEJ JAKO WYRAZ PRZYSTOSOWANIA DO FUNKCJI
Jednostkami budulcowymi tkanki nerwowej są - podobnie jak to ma miejsce w tkankach innego rodzaju - komórki. Komórki nerwowe, zwane neuronami lub neurocytami, różnią się pod względem strukturalnym w zasadniczy sposób od komórek innego typu (ryc.18).
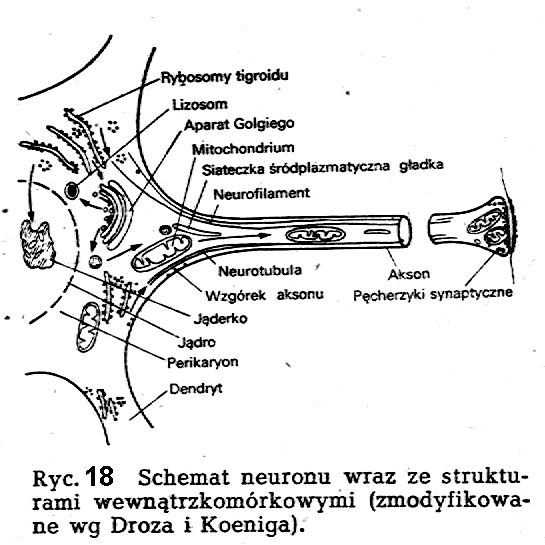
Ta
odmienność strukturalna-polegająca przede wszystkim na istnieniu
długich wypustek komórkowych -jest wyrazem przystosowania
neuronu do jego funkcji. W neuronie wyróżnić można następujące
cztery strefy czynnościowe związane z podstawowymi jego funkcjami: a)
wejście, b) inicjacja impulsów, c) przewodzenie impulsów
i d)
wyjście. Wymienione tu struktury neuronu mogą się wykształcić w rozmaity sposób, co jest podstawą klasyfikacji komórek nerwowych na różne typy strukturalni-czynnościowe.
Podstawowe struktury neuronu
Mimo że zarówno w różnych obszarach układu nerwowego u tego samego osobnika, jak też u zwierząt różnych gatunków - spotkać można neurony o rozmaitych kształtach, podstawowe struktury wszystkich neuronów są w zasadzie jednakowe (ryc.18). Pierwszym podstawowym elementem strukturalnym komórki nerwowej jest ciało neuronu (soma). Ciała neuronów mogą być rozmaitego kształtu: od okrągłych poprzez gwiaździste do wrzecionowatych. Ciało neuronu przechodzi w dwa typy wypustek: aksony (dawna nazwa: wypustki osiowe) i dendryty (dawna nazwa: wypustki protoplazmatyczne),
Akson, czyli neuryt, jest głównym rejonem przewodzenia w neuronie, a wiec główną drogą komunikowania się z inną komórką nerwową, lub też z komórką efektora (np. mięsień, gruczoł). Ze względu na spełniane funkcje akson, który jest pojedynczą wypustką neuronu, zwykle charakteryzuje się dużą długością. Akson bierze swój początek w obszarze neuronu zwanym odcinkiem początkowym aksonu. Jest to element inicjacji impulsów. Zakończenie aksonu, zwykle mniej lub bardziej rozgałęzione, tworzy element wyjścia neuronu.
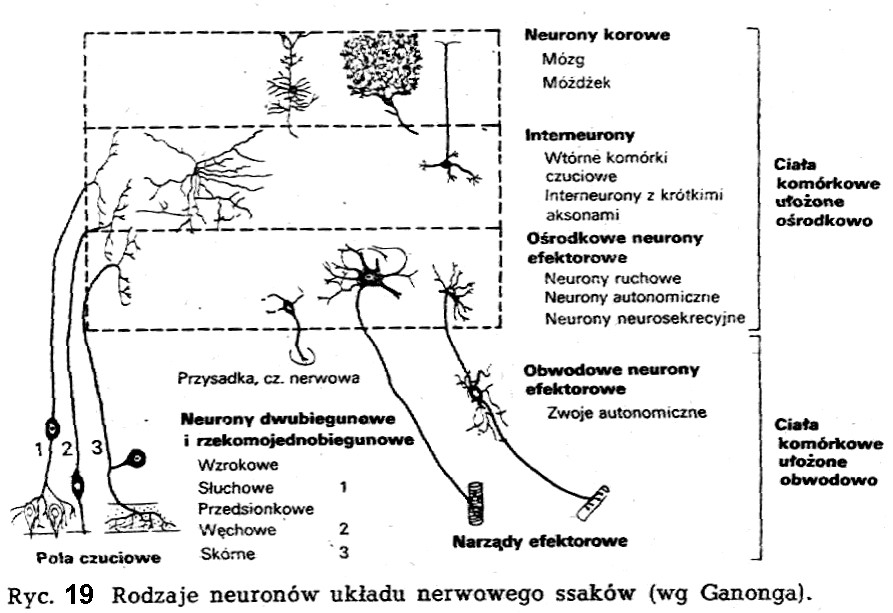
Ważnym
z punktu widzenia czynności neuronu elementem strukturalnym są
osłonki aksonu. W budowie struktur osłonowych uczestniczą komórki
należące do tkanki glejowej. W zależności od typów osłonek
okrywających akson, wyróżniamy włókna nerwowe z osłonką
mielinowa., czyli rdzenne, i włókna nerwowe bez osłonki
mielinowej, czyli bezrdzenne.
We włóknach rdzennych akson otoczony jest dwoma osłonkami: osłonką mielinową oraz neurolemą (dawna nazwa: osłonka Schwanna). Osłonka mielinową nie ma charakteru ciągłego. Przerywa się ona w regularnych odstępach, tworząc cieśni węzłów (dawna nazwa: węzły Ranyiera, przewężenia Ranviera). Neurolemą pokrywa osłonkę mielinową na przestrzeni całego włókna, a więc także w obrębie cieśni węzłów.
Drugim rodzajem wypustek neuronu są deadryty, stanowiące element wejścia neuronu. Występują one w neuronie w znacznej niekiedy liczbie i zwykle są krótsze od aksonu, a poza tym cechują się posiadaniem licznych drzewiastych rozgałęzień.
Spośród struktur specyficznych dla komórek nerwowych, które pojawiły
się w neuronach na skutek specjalizacji czynnościowej, należy wymienić: neurofibryle, tigroid (dawna nazwa: substancja Nissla) oraz wyspecjalizowaną w zakresie recepcji i przepuszczalności błonę komórkowa. Neurofibryle są jednym z najbardziej charakterystycznych tworów występujących w komórce nerwowej. Są to cieniutkie nitkowate twory, wnikające do wszystkich wypustek neuronu. Jednostką mniejszą od neurofibryli są neurofila-menty. Struktury te tworzą układ neu-rotubuli. Niegdyś przypisywano neuro-tubulom zasadniczą i dominującą role w przenoszeniu pobudzenia wzdłuż aksonu. Dzisiaj nie ulega wątpliwości, że układ neurotubuli obok funkcji budulcowej odgrywa ważną rolę w transporcie neurohormonów, czy też przekaźników nerwowych z ciała neuronu do zakończeń aksonu. Grudki substancji zasadochłonnej tworzące tigroid są rozsiane w całej cytoplazmie neuronu. Brak ich w aksonie; w dendrytach występują obficie. Tigroid, składający się z rybosomów i siateczki śródplazma-tycznej, jest skupiskiem kwasów nukleinowych i odgrywa ważną role w procesach przemiany materii i energii w neuronie.
POTENCJAŁY ELEKTRYCZNE KOMÓRKI NERWOWEJ
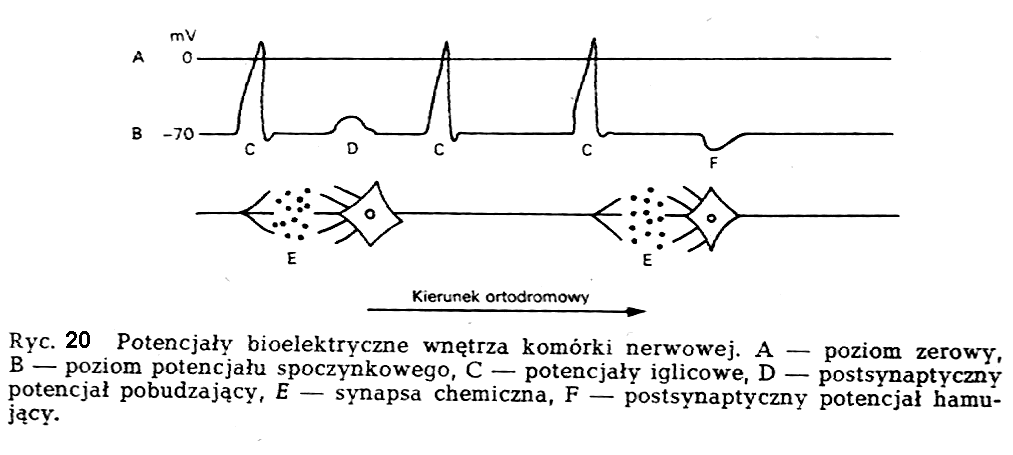
Jednym
z podstawowych przejawów życiowych komórki nerwowej
jest jej zdolność do generowania i przewodzenia potencjałów
elektrycznych. Te właściwości komórki nerwowej zlokalizowane
są przede wszystkim w błonie komórkowej, która oddziela
neuron od jego otoczenia.
Wśród potencjałów bioelektrycznych można wyróżnić potencjał spoczynkowy oraz potencjały czynnościowe. Potencjał spoczynkowy występuje, gdy neuron nie jest pobudzany. Potencjały czynnościowe towarzyszą stanowi czynnemu neuronu (ryc.20).
Metody badania potencjałów bioelektrycznych
Istotny postęp w badaniach, neurofizjo-logicznych szedł zawsze w parze z rozwojem technik badawczych. Wśród technik stosowanych, do badania neuonu na pierwsze miejsce wysuwają się metody rejestracji potencjałów bioelektrycznych. Każda z takich metod zawiera w sobie dwa zasadnicze elementy: element stykający się bezpośrednio ze źródłem generowania potencjałów bioelektrycznych (elektroda) oraz część rejestrującą (np. oscyloskop). Miedzy te dwa podstawowe elementy wyposażenia rejestrującego wprzęgnięte są zazwyczaj dodatkowe urządzenia w postaci np. przedwzmacniaczy.
W związku z tym, że neurony generujące potencjały bioelektryczne wytwarzają wokół siebie pole elektryczne, poza rejestracją z wnętrza komórki istnieje także możliwość rejestracji zewnątrzkomórkowej potencjałów bioelektrycznych.
Doświadczenia z wewnątrzkomórkowym pomiarem potencjałów bioelektrycznych w neuronie datują się od lat 1939-1940, kiedy to czterem uczonym (Cole, Curtis, Hodgkin, Huxley) udało się po raz pierwszy wprowadzić mikro-elektrodę do olbrzymiego aksonu mątwy, zwanego tak ze względu na średnicę dochodzącą do 1 mm. Podstawowe elementy techniki wewnątrzkomórkowej rejestracji potencjałów bioelektrycznych neuronu przedstawiono na ryc.21
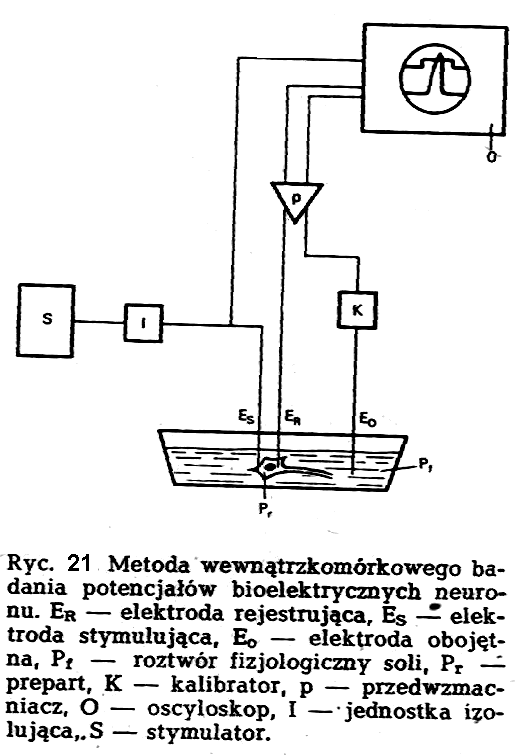
Obok
metod rejestracji potencjałów bioelektrycznych ważnym
elementem techniki badania komórki nerwowej jest metoda
stymulacji. W zakresie badań nad pojedynczą komórką nerwową
stosuje się obecnie przede wszystkim
stymulacją elektryczną i stymulacje chemiczną. Metoda dokom orkowego wprowadzania substancji chemicznych nosi nazwę mikroelektroforezy lub jontoforezy.
Potencjał spoczynkowy i jego geneza
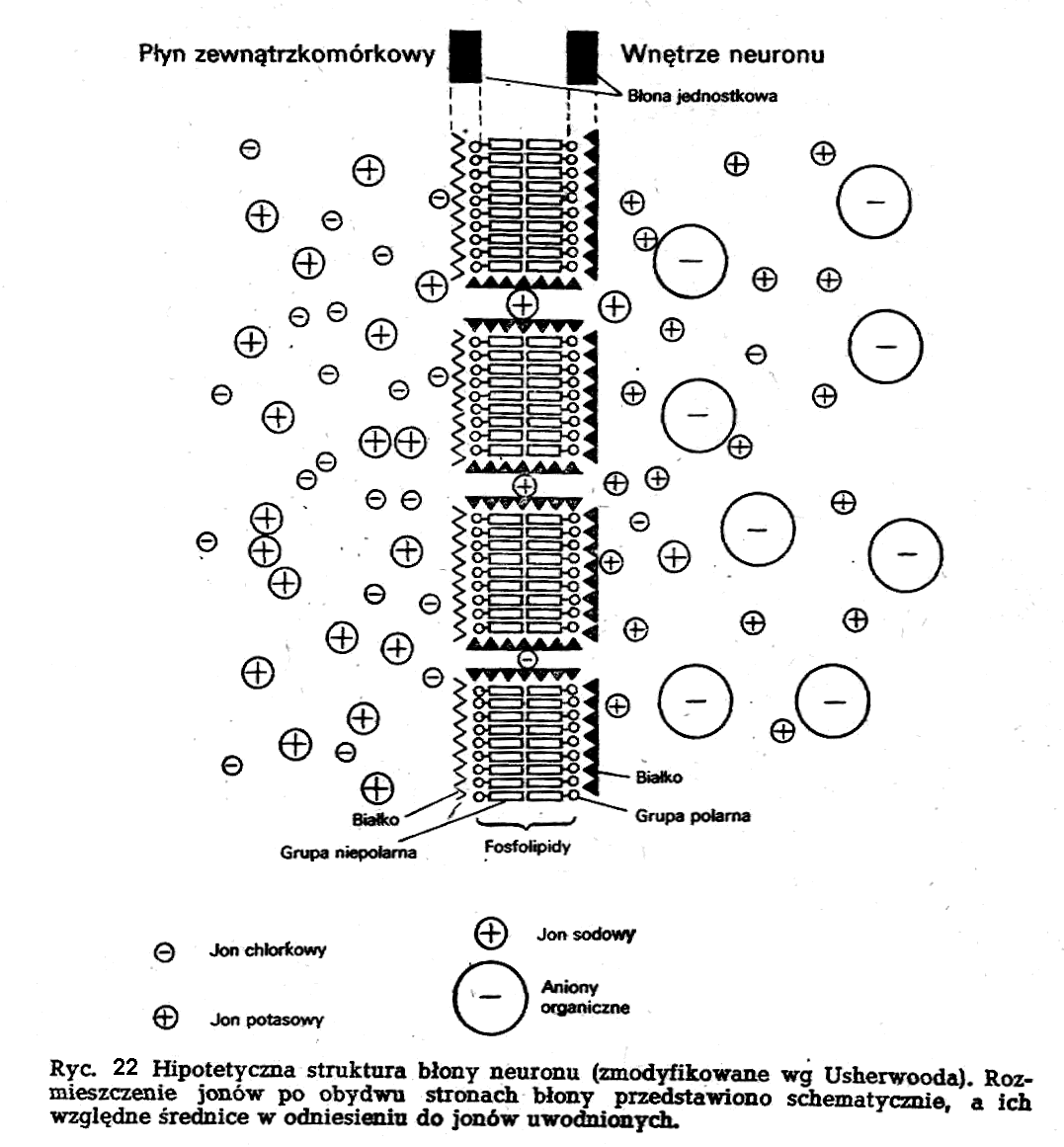
Powierzchnia nieuszkodzonego ciała neuronu, czy też jego włókien w spoczynku jest izopotencjalna. Znaczy to, że pomiędzy dwoma dowolnymi punktami na powierzchni neuronu nie ma różnicy potencjałów. Stwierdzana w neuronach w czasie spoczynku różnica potencjałów między wnętrzem a otoczeniem nosi nazwę potencjału spoczynkowego lub błonowego potencjału spoczynkowego. Różnica ta wynosi kilkadziesiąt miliwoltów.
Poza wieloma podobieństwami w składzie cytoplazmy neuronu i jego płynnego środowiska zewnętrznego istnieją istotne różnice w dystrybucji jonów w obydwu tych środowiskach (ryc. 22). W środowisku zewnątrzkomórko-wym głównym anionem jest chlor, którego stężenie jest tu około 10 razy wyższe niż w cytoplazmle neuronu. U źródeł tego nierównomiernego rozłożenia jonów chlorkowych leży przede wszystkim obecność w cytoplazmie dużych, nie przechodzących przez błonę anionów organicznych. W związku z tym można powiedzieć, że rozmieszczenie jonów chlorkowych w środowisku wewnątrz- i zewnątrzkomórkowym jest wynikiem równowagi Donnana.
Z jonów odgrywających pierwszorzędową rolę w genezie zjawisk bioelektrycznych należy wymienić sód i potas. Sód jest około 10 razy bardziej stężony na zewnątrz neuronu niż w jego wnętrzu. Stężenie natomiast potasu jest 30 razy wyższe w cytoplazmie neuronu niż w jego otoczeniu. Jeśli chodzi o sód, to biorąc pod uwagę ujemność wnętrza neuronu, iatwo dojść do wniosku, że siły wynikające zarówno z gradientu stężeń, jak i gradientu elektrycznego działają w tym samym kierunku (dokomórkowo). Obserwowane asymetrie w dystrybucji jonów sodowych i potasowych w środowisku zewnątrz i wewnątrzneuronalnym nie mogą być wynikiem biernych procesów błonowych. Asymetrie te są wynikiem aktywnego transportu jonów sodowych i potasowych. Transport ten dochodzi do skutku dzięki działalności w błonie neuronu systemu pomp sodowych i potasowych.
U podstaw tych układów leży działalność specjalnego błonowego enzymu transportującego - adenozynotrójios-fatazy (Na-K-ATP-aza), aktywowanej przez Na+ i K+. Proces transportu jonów przebiega ze zużyciem adenozyno-trójfosforanu (ATP). W następstwie hydrolizy ATP powstaje adenozynodwu-fosforan (ADP) i fosforan nieorganiczny. W czasie powyższej reakcji wyzwala się energia pochodząca z hydrolizy bogatego energetycznie wiązania fosforanowego. Energia ta zużywana jest do transportu jonów wbrew gradientowi stężeń. Szereg cech czynnościowych Na-K-ATP-azy pokrywa się z cechami pompy j ono we j, wskazuj ąc na ścisiy związek tych dwóch mechanizmów.
Częściowe lub całkowite zahamowanie działania pompy sodowo-potasowej nastąpić może poprzez: 1) niedotlenienie spowodowane brakiem tlenu lub działaniem inhibitorów oddychania komórkowego, 2) zastosowanie wybiórczych inhibitorów pompy, np. ouabainy i pokrewnych glikozydów" naserco-wych, 3) spadek temperatury, powodujący obniżenie metabolizmu komórkowego.
Na skutek aktywności transportującej ATP-azy jony sodowe nieustannie usuwane są na zewnątrz komórki, a do jej wnętrza wprowadzane są jony potasowe, przy czym stosunek wymiany wynosi dwa jony potasowe za trzy jony sodowe. Transportująca ATP-aza aktywowana jest przez jony sodowe działające na wewnętrzną powierzchnie komórki. Im większy jest wobec tego napływ tych jonów do wnętrza na skutek biernej dyfuzji, tym większa je^t aktywność pompy jonowej i tym większy czynny transport na zewnątrz. Na skutek istnienia tego ujemnego sprzężenia zwrotnego czynny transport sodu jest równy biernej dyfuzji tych jonów, wobec czego średni ich przepływ przez błonę pozostającą w stanie spoczynku równa się 0.
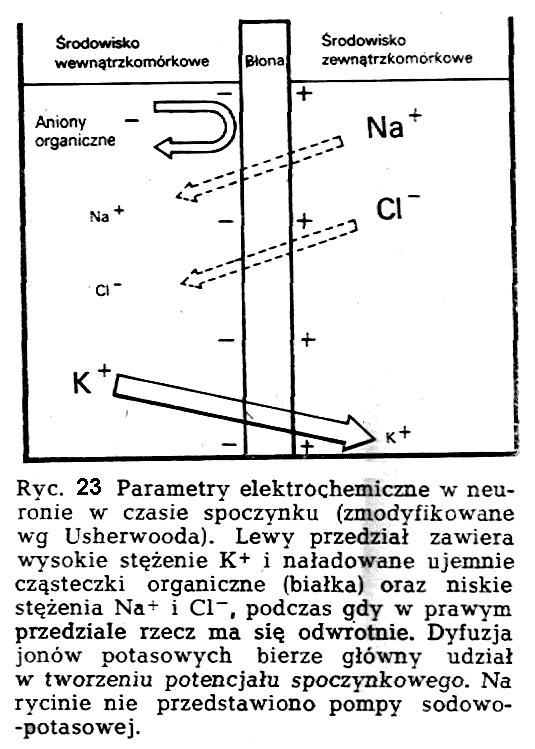
W
poprzek błony neuronu istnieją wyraźne gradienty stężeń dla jonów
sodowych, potasowych i chlorkowych (ryć. 23). Pozostająca w spoczynku
błona neuronu jest wysoce przepuszczalna dla jonów potasowych,
mniej dla chlorkowych i minimalnie dla jonów sodowych. Jeżeli
przepuszczalność błony neuronu dla jonów potasowych
przyjmujemy jako jedność, to przepuszczalność dla omawianych trzech
jonów (K+, CI-, Na+) wynosi odpowiednio-1 : 0,45 : 0,04.
Biorąc to pod uwagę, a także pamiętając, że istnieje asymetria w
stężeniach jonów w poprzek błony, łatwo dojść do wniosku, że w
poprzek błony neuronu musi istnieć różnica potencjałów.
Te różnicę potencjałów określamy, jako potencjał
spoczynkowy. Wielkość błonowego potencjału spoczynkowego określana
jest głównie przez wielkość gradientu stężeń dla jonów
potasowych i zbliża się do potencjału równowagi dla potasu.
Tak rozumianą spoczynkową różnice potenciałów wyraża się wzorem:
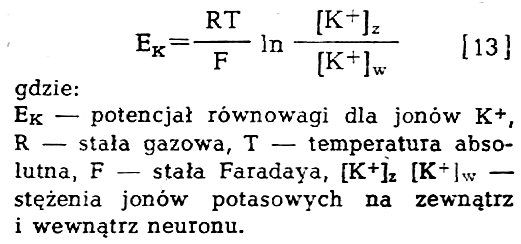
Z
wzoru [13] łatwo wyciągnąć wniosek, że zwiększenie stężenia potasu w
środowisku zewnątrz neuronu prowadzić musi do zmniejszania różnicy
potencjału w poprzek błony neuronu. Należy pamiętać, że w omawianym
przypadku mamy do czynienia z neuronem zachowującym się jak elektroda
potasowa; nie można jednak wykluczyć udziału innych jonów w
kształtowaniu spoczynkowej różnicy potencjałów. Ich
rola jest jednak minimalna.
Ponieważ prawidłowy przebieg potencjałów czynnościowych i wszystkich. zjawisk z nimi związanych, a przede wszystkim przewodnictwa stanu czynnego, zależy od potencjału spoczynkowego, na tle którego komórki zostały aktywowane, prawidłowa czynność komórek zależy w dużym stopniu właśnie od prawidłowego potencjału spoczynkowego.
W stanach chorobowych przebiegających ze znacznego stopnia hiperpota-semią, tj. zwiększeniem stężenia jonów potasowych we krwi i płynach zewnątrz-komórkowych, dochodzi do bardzo poważnych zakłóceń czynności serca, którego komórki posiadają właściwości elektrofizjologiczne zbliżone do właściwości neuronów. Wzrost stężeń potasu zewnątrzkomórkowego, zgodnie ze wzorem Nernsta, powoduje spadek potencjału spoczynkowego, będącego, jak już wspomniano, potencjałem równowagi dla jonów potasowych. Powstające na tle obniżonego potencjału spoczynkowego potencjały czynnościowe przebiegają nieprawidłowo, co powoduje zaburzenia w przewodzeniu stanu czynnego i w pobudliwości.
Potencjał czynnościowy i jego geneza
Istnienie potencjału spoczynkowego jest nieodzownym warunkiem dla powstania czynnościowych zjawisk bioelektrycznych w neuronie. Punktem wyjścia dla zjawisk czynnościowych jest zmiana stanu spolaryzowania neuronu. Zmiana ta może iść w kierunku zmniejszenia spoczynkowej różnicy potencjałów, wtedy nosi nazwą depolaryzacji, lub też polegać może na zwiększeniu stanu spolaryzowania i nazywana jest hiperpolaryzacją.
Bardzo pouczające jest prześledzenie poszczególnych etapów powstawania potencjału czynnościowego. Jeżeli na neuron stosuje sią elektryczny bodziec depolaryzacyjny, wtedy na oscyloskopowym zapisie wewnątrzkomórkowym zaczną się pojawiać zmiany świadczące o depolaryzacji błony komórki, a więc o zmniejszaniu się potencjału spoczynkowego. Zmiany te, nazywane elektro-tonicznymi, mają charakter bierny i są odzwierciedleniem specyficznej budowy błony neuronu, która nosi w sobie cechy kondensatora. W dalszym ciągu na szczycie zmian elektrotonicznych zaczyna się pojawiać zmiana potencjału, .nazywana odpowiedzią lokalną, która jest już czynną odpowiedzią neuronu na bodziec depolaryzacyjny, choć nie posiada właściwości rozprzestrzeniania się. Gdy opisywane tu zmiany osiągną poziom depolaryzacji krytycznej lub progowej, dochodzi w neuronie do powstania właściwego potencjału czynnościowego, nazwanego ze względu na kształt potencjałem iglicowym (ryć. 24). Jest to potencjał pojawiający sią zgodnie z prawem "wszystko albo nic". Oznacza to, że gdy na skutek stymulacji zmiany elektrotoniczne osiągną w neuronie poziom depolaryzacji krytycznej i powstanie potencjał iglicowy, dalsze pobudzanie nie zmienia w danych warunkach amplitudy powstałego potencjału. Istotną cechą potencjału czynnościowego neuronu jest zdolność do rozprzestrzeniania się.
Na ryc. 24 przedstawiono omówione fazy powstawania potencjału czynnościowego oraz jego strukturę. Za główne mechanizmy spoczynkowej różnicy potencjałów uważa się gradienty stężeń jonów po obydwu stronach błony neuronu, przepuszczalność błony dla tych jonów oraz pompę Na+ - K+. Jeżeli przyjmuje się, że potencjał czynnościowy jest zaburzeniem stanu spoczynkowego, musi dojść do zmian wymienionych wyżej parametrów. Neuron nie może zmienić stężeń jonowych z szybkością i w rozmiarach, które by w istotny sposób wpłynęły na wielkość potencjału błonowego. Neuron jest jednak w stanie- zmienić właściwości swej błony, a szczególnie przepuszczalność dla jonów. Tak też jest w istocie.
W
myśl teorii zaproponowanej przez A. L. Hodgkina i A. F. Huxleya
(1939), potencjał czynnościowy neuronu powstaje w wyniku nagłego,
dużego, przejściowego wzrostu przepuszczalności błony neuronów
dla Na+. Przez bardzo krótki okres przepuszczalność błony
neuronu dla Na+ przewyższa przepuszczalność błony dla innych jonów.
Z badań doświadczalnych wynika, że w czasie potencjału czynnościowego
przepuszczalność błony neuronu dla Na+ wzrasta kilkaset razy w
porównaniu że stanem spoczynkowym. Jony Na+, wchodząc zgodnie
z gradientem stężeń do komórki zmieniają potencjał wnętrza
neuronu z ujemnego na dodatni (ryc.25). Podwyższenie przepuszny
potencjał czynnościowy. Jest to zjawisko refrakcji.
Wkrótce
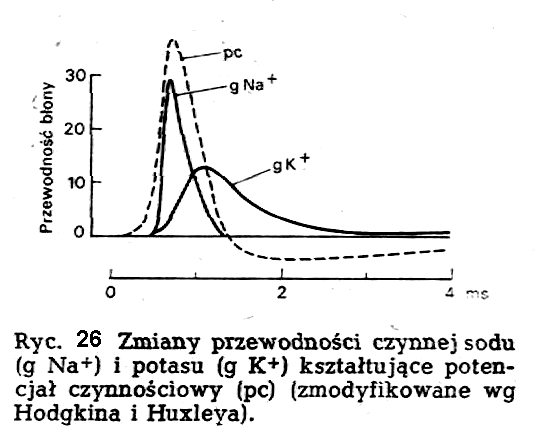
po rozpoczęciu aktywacji Na+dochodzi także do wzrostu
przepuszczalności błony dla jonów K+. Proces ten jest nazywany
aktywacją potasową. Szybkość narastania wzrostu przepuszczalności dla
jonów K+ jest znacznie mniejsza niż dla jonów Na+.
Aktywacja K+ osiąga swoje maksimum w chwili, gdy przepuszczalność
błony dla sodu powraca do wartości wyjściowych (in-aktywacja sodowa).
Przepuszczalność błony neuronu dla jonów Na+ jest największa
na szczycie potencjału czynnościowego, po czym ulega szybkiemu
zmniejszeniu. W tym obszarze różnica potencjału w poprzek
błony neuronu zbliża się do potencjału równowagi dla )Na+
(ENa). Z tego powodu w wielu neuronach szczyt potencjału
iglicowego można przewidzieć na podstawie stężeń sodu na zewnątrz
[Na+]z i wewnątrz [Na+]w neuronu w myśl równania Nernsta
zastosowanego do sodu
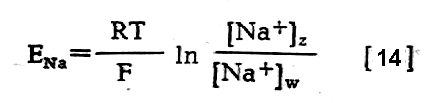
Proces
inaktywacji sodowej i aktywacji potasowej jest odpowiedzialny za
repo-laryzację neuronu. Repolaryzacja kończy potencjał czynnościowy.
Gdy potencjał równowagi dla potasu nie pokrywa się dokładnie z
potencjałem spoczynkowym, wtedy po fazie repolary-zacji przez kilka
milisekund potencjał błonowy będzie różnił się od potencjału
spoczynkowego. Te przejściowe zjawiska noszą nazwę potencjałów
następczych; mogą one mieć charakter 'zarówno depolaryzacyjny,
jak i hiperpola-ryzacyjny (ryc. 26). Z przytoczonego opisu genezy
iglicowego potencjału czynnościowego wynika, że dla powstania tego
potencjału nieodzowna jest obecność jonów Na+ w środowisku
otaczającym neuron.
Wnikliwa
analiza opisanych powyżej procesów jonowych, biorących udział
w genezie potencjału czynnościowego, stała się możliwa dzięki
zastosowaniu nowoczesnej metody stabilizacji potencjału komórkowego
(voltage. clamp). Metoda ta pozwala na rozdzielenie i
zidentyfikowanie prądów jonowych niesionych przez jony Na+ i
K+ w różnych fazach powstawania potencjału czynnościowego.
Okazało się np., że potencjał krytyczny (progowy) błony komórkowej
neuronu jest momentem, w którym prąd sodowy jest równy
prądowi potasowemu (przy odwrotnych kierunkach).
Zastosowanie nowoczesnych metod badawczych pozwoliło na opracowanie hipotetycznej koncepcji tzw. kanałów w błonie neuronu, a więc miejsc, w których zlokalizowane są procesy aktywacji i inaktywacji jonowej. Istnieje szereg koncepcji strukturalnych kanałów błonowych. Właściwości kanału związane z przepuszczalnością jonów przez błoną wiążą się z uruchomieniem, podtrzymywaniem czy wreszcie zahamowaniem przechodzenia jonów przez dany kanał. Hipotetyczny kanał błony uważany jest także za mechanizm posiadający zdolność wyboru jonów. W związku z tym istnieją przypuszczenia o istnieniu np. odrębnych kanałów sodowych czy też potasowych.
Punktem przełomowym w badaniach nad mechanizmami aktywacji i inaktywacji jonowej w czasie potencjału czynnościowego było wprowadzenie do tych badań tetrodotoksyny. Jest to jad produkowany przez żyjące głównie w Oceanie Spokojnym ryby z rodziny Te-trodontidae. Jad ten posiada zdolność
wybiórczego blokowania wzrostu przepuszczalności błony neuronu dla jonów Na+; eliminuje to możliwość powstania potencjału czynnościowego. W związku z tym tetrodotoksyna jest jedna z najsilniejszych trucizn. Stosując ją ustalono, że blokowanie wzrostu przepuszczalności błony dla Na+ polega na łączeniu się jednej cząsteczki tetrodotoksyny z jednym kanałem sodowym. Doświadczenia te pozwoliły na ustalenie liczby miejsc (kanałów) sodowych w błonie neuronu. Liczbę tę szacuje się średnio na około 50 kanałów na 1mm2 błony.
POBUDLIWOŚĆ, POBUDZENIE l HAMOWANIE
Pobudliwość można określić jako zdolność organizmów, tkanek, czy też komórek do odpowiadania na bodźce. Wykazujące tę zdolność organizmy, tkanki, czy też komórki nazywamy pobudliwymi. Różne organizmy, a w danych organizmach określone tkanki czy też komórki, posiadać mogą różną pobudliwość. Niekiedy te same elementy mogą wykazywać różnice pobudliwości w czasie; w ramach rytmiki biologicznej można obserwować zmiany pobudliwości w ciągu np. doby, miesiąca, czy tez roku. W kategoriach całego organizmu odpowiedzi realizowane są jako wynik działania bodźców na specjalnie do tego przystosowane elementy - receptory, będące podstawowym elementem narządów zmysłów. Na poziomie komórki bodźce działają na błonę komórkową.
Określenie poziomu pobudliwości organizmu, tkanek, czy też komórek dokonuje się przede wszystkim metodą pośrednią. Metoda ta polega na pomiarze wielkości stosowanego bodźca. Bardzo pożyteczną, a zarazem powszechnie stosowaną, miarą poziomu pobudliwości jest próg pobudliwości. Przez próg pobudliwości rozumie się najsłabszy bodziec zdolny do wywołania w danych warunkach określonej reakcji. Jest to bodziec progowy. Bodźce o natężeniu niższym od progowego nazywamy pod-progowymi, o natężeniu wyższym zaś bodźcami nadprogowymi. Ponieważ na organizm, tkankę, czy też komórkę działać mogą bodźce niosące w sobie różne rodzaje energii, bodźce progowe - a tym samym także próg pobudliwości - mogą posiadać różne miana. Można wiec wyrażać próg pobudliwości w miliwoitach lub miliamperach (bodziec elektryczny prądów selektywnych), milimolach (bodziec chemiczny) itd.
Jak wynika z przytoczonej tu definicji, pobudliwość przejawia się w zdolność: odpowiadania na bodźce. Komórka nerwowa może odpowiadać na bodźce w rozmaity sposób (np. zmianami częstotliwości potencjałów czynnościowych). Jeżeli jednak jako stan wyjściowy przyjmie się neuron w stanie spoczynku, to na działanie bodźca może on zareagować w dwojaki sposób: zmniejszeniem spoczynkowej różnicy potencjału (depolaryzacją) lub jej zwiększeniem (hiperpolaryzacją). W pierwszym przypadku dochodzi do pobudzenia neuronu, w drugim zaś do jego zahamowania, co można określić z jednej strony jako stan uczynnienia, z drugiej zaś strony jako unieczynnienie neuronu. Zarówno depolaryzacja, jak i hiperpola-ryzacja są procesami aktywnymi) wymagającymi energii.
Przewodzenie informacji w obrębie neuronu
W neuronie bardzo jaskrawo uwidocznione są powiązania i zależności strukturalno-czynnościowe. Wyodrębniono w strukturze neuronu cztery zasadnicze strefy czynnościowe: 1) wejście neuronu (dendryty i częściowo ciało neuronu), 2) strefa inicjacji impulsów (początkowy odcinek akso-nu). 3) strefa przewodzenia (głównie akson), 4) wyjście neuronu (zakończenie aksonu). Kolejność, w jakiej podano strefy czynnościowe, jest zarazem obrazem kolejności uczynniania poszczególnych elementów neuronu, a także kierunku przewodzenia informacji w obrębie neuronu.
Dendryty, stanowiące jeden z elementów wejściowych neuronu, charakteryzują się określonymi cechami zarówno strukturalnymi, jak i czynnościowymi, odróżniającymi je od pozostałych części neuronu. W ramach cech strukturalnych na uwagę zasługuje tzw. aparat pączkowy (spine apparatus), znajdujący się w "pączkach" ("kolcach") dendrytów. Aparat składa się z równoległych warstw pęcherzyków. Strukturze tej przypisuje się udział w procesach związanych z torowaniem. Z cech czynnościowych obszaru dendrytycznego wymienić należy znacznie wyższy próg pobudliwości w porównaniu z początkowym odcinkiem aksonu oraz znacznie mniejszą prędkość przewodzenia impulsów w porównaniu z prędkościami występującymi w aksonach.
Przewodzenie informacji od dendrytów do zakończeń aksonu jest kierunkiem fizjologicznym i nosi nazwę przewodzenia ortodromowego. Kierunek przeciwny (nie występujący w organizmach żywych, lecz możliwy do uzyskania eksperymentalnie) nosi nazwę przewodzenia antydromowego.
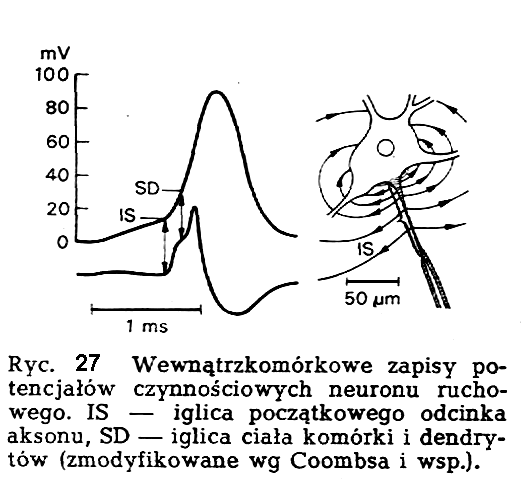
Na ryc. 27 przedstawiono wewnątrzkomórkowe zapisy zjawisk bioelektrycznych w neuronie ruchowym. Z zapisów tych widać, że ramię wstępujące potencjału iglicowego wykazuje w swym przebiegu załamki. Jeśli ten charakterystyczny przebieg podda się szczegółowej analizie poprzez zastosowanie różniczkowania elektrycznego, wtedy wyraźnie widać, że na właściwy potencjał czynnościowy składają się oddzielone od siebie dwa elementy bioelektryczne. Te dwa elementy są obecnie określane ze względu na miejsce powstawania w neuronie jako iglica odcinka początkowego aksonu wraz ze wzgórkiem aksonu oraz iglica ciała komórki i dendrytów. Badania elektrofizjo-logiczne wykazały istnienie różnic w poziomie depolaryzacji krytycznej w rejonach odcinka początkowego aksonu oraz ciała komórki i dendrytów. Jeśli chodzi o dotychczas zbadane neurony ruchowe, to próg odcinka początkowego aksonu jest zawsze niższy od progu ciała komórki i dendrytów i co więcej, stosunek ten wynosi co najmniej 1 : 2 tzn., że próg ciała komórki i dendrytów jest co najmniej dwukrotnie wyższy od progu odcinka początkowego aksonu. Stymulacja wejścia neuronu (dendryty i ciało neuronu) nie prowadzi bezpośrednio do powstania potencjału czynnościowego w tym rejonie, a jedynie do szerzenia się fali depolaryzacji w kierunku odcinka początkowego aksonu, który ze względu na niski próg jest miejscem wyzwolenia potencjału czynnościowego (ryc.27).
Główną
strefą przewodzenia fali depolaryzacji w neuronie jest akson. W
przewodzeniu aksonalnym zaznaczają się wyraźnie ścisłe związki między
strukturą a funkcją. Po wyzwoleniu we włóknie nerwowym
potencjału czynnościowego w miejscu wyzwolenia nie może w okresie ok.
l ms powstać następna odpowiedź. Okres, w którym nie może
powstać - bez względu na natężenie stosowanego bodźca - kolejny
potencjał czynnościowy, nazywany jest okresem refrakcji bezwzględnej.
W przebiegu powstawania potencjału czynnościowego okres refrakcji bezwzględnej odpowiada okresowi inakty-wacji sodowej, a wiać -stopniowego zmniejszania przepuszczalności błony dla jonów Na+. Po okresie refrakcji bezwzględnej następuje okres, w którym neuron odzyskuje wprawdzie swą pobudliwość, lecz próg depolaryzacji krytycznej jest podwyższony (do wywołania potencjału czynnościowego konieczny jest silniejszy bodziec). Ten okres trwający do kilku milisekund (w zależności od rodzaju włókien) nazywany jest okresem refrakcji względnej. Zjawisko refrakcji przeciwdziała nakładaniu się potencjałów czynnościowych na siebie; w związku z tym występują one jako odrębne zjawiska bioelektryczne. Należy także zdać sobie sprawę z tego, że okresy refrakcji determinują w dużej mierze częstotliwości wyładowań neuronu.
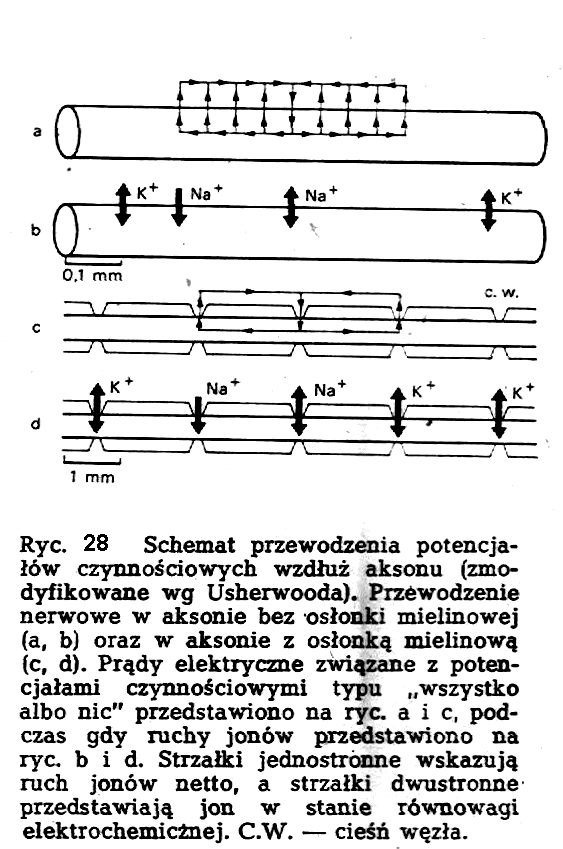
Sposób
przewodzenia informacji we włóknach nerwowych zależy głównie
od tego, czy włókna te msją osłonkę mielinową, czy też są jej
pozbawione. We włóknach bez osłonki mielinowej potencjały
czynnościowe wędrują ruchem jednostajnym ze stałą dla danych warunków
prędkością. Ten typ przewodzenia nosi nazwę przewodzenia ciągłego. We
włóknach z osłonką mielinową
przewodzenie potencjałów czynnościowych odbywa się z niejednolitą prędkością. Prędkość przewodzenia jest bardzo duża w obszarach pomiędzy ciemniami węzłów, natomiast w samych cie-śniach węzłów następuje "przestój" potencjału czynnościowego. Ten rodzaj przewodzenia nazywa się przewodzeniem skokowym. Zasady działania mechanizmów przewodzenia ciągłego i skokowego obrazuje ryc. 28.
Ze względu na stałe procesy odnowy jonowej (w przewodzeniu ciągłym wzdłuż całego włókna, a w przewodzeniu skokowym w cieśniach węzłów) amplituda potencjału czynnościowego nie maleje w miarę przesuwania się tego potencjału wzdłuż włókien nerwowych. Jest to zjawisko zwane przewodzeniem bez dekrementu (strat). W niektórych przypadkach, szczególnie przy zmianach chorobowych, występuje przewodzenie z dekrementem, co oczywiście zaburza czynność układu nerwowego.
Prędkość przewodzenia impulsów wzdłuż włókien nerwowych zależy od średnicy włókien nerwowych oraz od sposobu przewodzenia. Prędkość przewodzenia potencjału czynnościowego zależy od podłużnej oporności włókna nerwowego. Ta oporność, podobnie jak w układach nieożywionych, jest odwrotnie proporcjonalna do kwadratu średnicy przewodnika. Wynika stąd, że włókna o dużym przekroju będą miały względnie niską oporność aksoplazmy i co za tym idzie - zdolność do większej prędkości przewodzenia potencjałów czynnościowych, włókna zaś o przekroju mniejszym będą miały wyższą oporność aksoplazmy i przewodzić będą wolniej. Obok wyraźnego wpływu wielkości przekroju włókna na prędkość przewodzenia fali depolaryzacji, ważnym czynnikiem ją kształtującym jest sposób przewodzenia potencjałów. We włóknach rdzennych, dzięki dobrym właściwościom izolacyjnym osłonki mielinowej, prędkość przewodzenia jest znacznie wyższa, niż wskazywałby na to porównawczo przekrój włókna. Prędkość przewodzenia fali depolaryzacji we włóknach nerwowych jest "dopasowana" do funkcji danych włókien.
Grapy włókien nerwowych
|
Grupa |
A |
B |
C |
||||
|
Podgrupa |
a |
b |
g |
d |
- |
s |
d.r. |
|
Średnica aksonu w mm |
12-20
|
5-12
|
3-6 |
2-5 |
+/-3 |
0,3-1,3 |
0,4-1,2 |
|
Prędkość przewodzenia w m/s |
70-120 |
30-70 |
15-30 |
12-30 |
3-15 |
0,7-2,3 |
0,5-2,0 |
|
Osłonka mielinowa |
+ |
+ |
+ |
+ |
+ |
- |
- |
|
Włókna nerwowe aferentne |
+ |
+ |
|
+ |
|
|
+ |
|
Włókna nerwowe eferentne |
+ |
|
+ |
|
+ |
+ |
|
|
Czas trwania potencjału czynnościowego w ms |
0,4-0,5 |
1,2 |
2 |
2 |
|||
|
Okres refrakcji bezwzględnej w ms |
0,4-1,0 |
1,2 |
2 |
2 |
|||
a więc do biologicznego znaczenia szybkości reakcji.
Na podstawie cech zarówno morfologicznych, jak i czynnościowych dzieli się włókna nerwowe na cztery następujące grupy:
1)Włókna nerwowe grupy A, do których należą włókna z osłonką mielinową, spełniające rolę zarówno aferentną (dośrodkową), jak i eferentną (odśrodkową). W grupie tej - biorąc pod uwagę średnicę włókien - wyróżnia się cztery podgrupy (a,b,g i d).
2)Włókna grupy B są częścią układu autonomicznego. Posiadają one osłonką mielinową i są włóknami przywspółczulnymi oraz współczulnymi przedzwojowymi.
3)Włókna grupy Cs (sympathetic C fibers) - to włókna wspóiczulne zazwojowe, pozbawione osłonki mielinowej.
4)Włókna grupy C d.r. (dorsal root C fibers) - to włókna wstępujące do rdzenia przez korzenie grzbietowe. Są one pozbawione osłonki mielinowej. Szczegółowe dane dotyczące powyższych grup włókien przedstawiono w tabeli
Przekazywanie informacji innym komórkom
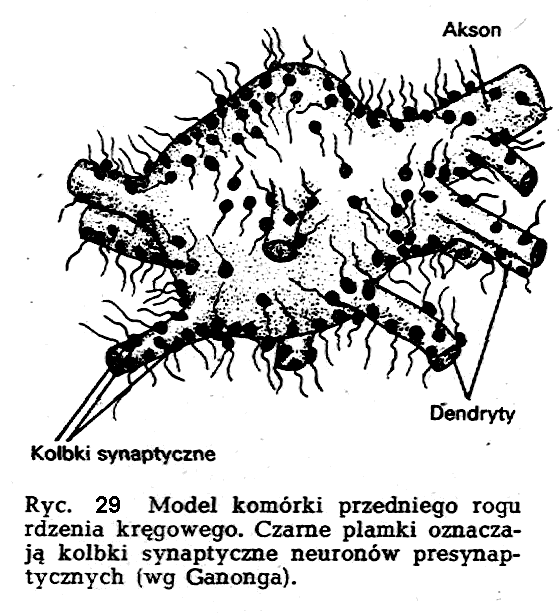
Informacja nerwowa, która dociera do wyjścia neuronu, a więc do zakończeń akscnów, musi być, zgodnie z założeniami czynnościowymi układu nerwowego, przekazana innym komórkom nerwowym, lub też efektorom, (np. mięśnie, gruczoły). Istnienie tej ciągłości czynnościowej jest nieodzownym warunkiem prawidłowego funkcjonowania układu nerwowego, a tym samym, oczywiście, całego organizmu. W parze ze wspomnianą ciągłością czynnościową nie idzie ciągłość strukturalna. Poszczególne komórki nerwowe nie są ze sobą zespolone, a jedynie stykają się ze sobą. To samo można powiedzieć o sposobie łączenia się zakończeń aksonu z efektorem. Ten obszar styku nosi nazwę synapsy. W zależności od rodzaju stykających się elementów wyróżnia się synapsy nerwowo-nerwowe (styk dwóch neuronów), synapsy nerwowo-mięśniowe. Połączenia między dwiema komórkami nerwowymi mogą przebiegać rozmaicie i w związku z tym wyróżnia się szereg synaps, których nazwy wskazują na charakter styku międzyneuronalnego. Wyróżnia się więc'synapsy aksodendrytyczne, aksoaksonalner aksosomatyczne (ryc. 29).
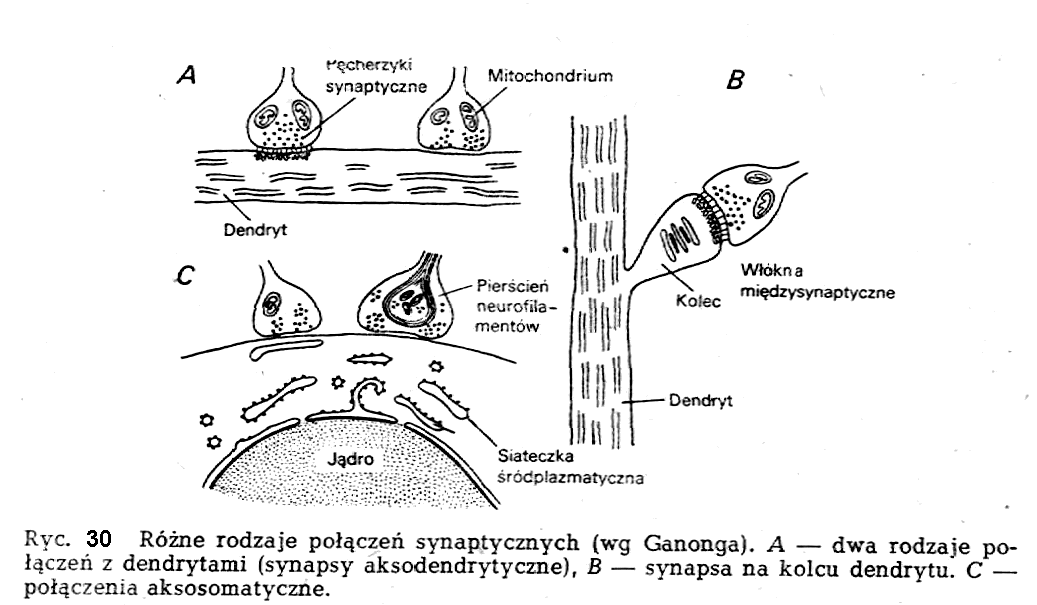
W
skład synapsy wchodzą trzy zasadnicze elementy strukturalne: element
presynaptyczny, szczelina synaptyczna i element postsynaptyczny.
Element presynaptyczny tworzą zakończenia aksonów. Błona
zakończeń jest nazywana błoną presynaptyczną. Szczelina synaptyczna
jest przerwą strukturalną między elementami prę- i
postsynap-tycznymi. W skład elementu postsynap-tycznego w synapsie
między dwoma neuronami mogą wchodzić różne części neuronu
postsynaptycznego (np. dendryty, ciało neuronu), w zależności od
charakteru synapsy. W związku z lokalizacją kluczowych procesów
postsy-naptycznych w błonie elementu postsynaptycznego wprowadzono
pojęcie błony postsynaptycznej.
Najbardziej istotnym momentem w przekazywaniu informacji innym komórkom poprzez synapsy jest zmiana nośnika dla informacji. W elemencie presynaptycznym nośnikiem dla przesyłania informacji są potencjały czynnościowe. W obrębie natomiast samej synapsy dochodzi do zmiany nośnika elektrycznego na chemiczny. Dzieje się to na skutek wydzielania przez element presynaptyczny substancji chemicznych zwanych mediatorami synaptycznymi (przekaźniki synaptyczne, substancje przekaźnikowe}. W obrębie synapsy właśnie te substancje przekazują nadaną przez element presynaptyczny informację nerwową do elementu postsynaptycznego. Synapsy posługujące się takim właśnie sposobem przekazywania informacji nazywane są synapsami chemicznymi.
Rola synaps i mediatorów chemicznych
Dochodzący - zgodnie z kierunkiem ortodromowym - do zakończeń akso-nu potencjał czynnościowy jest mechanizmem spustowym dla zapoczątkowania procesu przenoszenia synaptycznego. Pierwszym etapem tego procesu jest tzw. sprzężenie elektrowydzielnicze. Jest to proces polegający na zainicjowaniu przez falę depolaryzacji uwalniania mediatora synaptycznego do szczeliny synaptycznej. Dla zabezpieczenia ciągłości czynnościowej w obrębie układu nerwowego konieczne jest przejście mediatora poprzez szczelinę synaptyczną i połączenie z elementem postsynaptycznym. To połączenie odbywa się na poziomie błony postsynap-tycznej, a ściślej, na poziomie receptora błonowego, specyficznego dla danego mediatora synaptycznego. W błonie postsynaptycznej ma miejsce sprzężenie cherniczno-elektryczne. Oznacza to, że informacja znów korzystać będzie z nośnika elektrycznego, W następstwie połączenia się mediatora synaptycznego z receptorem^ błonowym powstają zmiany przepuszczalności błony postsynaptycznej dla jonów. Wynikiem tych zmian jest powstanie zjawisk bioelektrycznych, które ogólnie nazywamy potencjałami postsynaptycznymi. Potencjały postsynaptyczne należą do grupy potencjałów nie rozprzestrzeniających się, a także stopniowanych. Ta ostatnia cecha, która jest przeciwieństwem mechanizmu typu "wszystko albo nic", oznacza, że amplituda potencjału zależy od natężenia bodźca. Potencjały postsynaptyczne mogą mieć charakter depolaryzacyjny, lub też hiperpolaryzacyjny. W pierwszym przypadku mogą one doprowadzić do depolaryzacji krytycznej i w rezultacie do wyzwolenia potencjału czynnościowego i dlatego mają charakter pobudzający. W związku z tym depolaryzacyjne potencjały postsynaptyczne nazywa się postsynap-tycznymi potencjałami pobudzającym: (EPSP - excitatory postsynaptic po-tentials). Postsynaptyczne potencjały hiperpolaryzacyjne poprzez zwiększę-' nie spoczynkowej różnicy potencjałów wpływają w sposób hamujący na błonę postsynaptyczną. Nazywa się je dlatego postsynaptycznymi potencjałami hamującymi (IPSP - inhibitory postsynaptic potentials). Te dwa rodzaje wyładowań postsynaptycznych decydują o przynależności danej synapsy do kategorii pobudzających, czy też hamujących.
W związku z procesami synaptycznymi wypływa problem pobudliwości błony neuronu w odniesieniu do specyficznych bodźców. Cała nieomalże powierzchnia błony neuronu wykazuje pobudliwość elektryczną. Ta pobudliwość nie jest jednakowa we wszystkich częściach neuronu. Najwyższą pobudliwość elektryczną (najniższy próg pobudliwości) wykazuje odcinek początkowy aksonu. Istnieją wszakże obszary błony neuronu, które należy uznać za niepobudliwe elektrycznie. Są to te obszary błony neuronu postsynaptycznego, które wchodzą w skład synapsy chemicznej. Nie odpowiadają one na bodźce elektryczne, natomiast wykazują pobudliwość chemiczną (zdolność do reakcji na działanie mediatora synaptycznego).
Innym parametrem związanym z czynnością synaps jest tzw. opóźnienie synaptyczne. Jest to czas, jaki jest potrzebny na przejście informacji nerwowej poprzez synapsę. W zależności od rodzaju synapsy i warunków, opóźnienie synaptyczne może być zawarte w granicach od 0,5 do kilku milisekund. Sama nazwa ,,opóźnienie" sugeruje, że synapsa opóźnia, .zwalnia przewodzenie informacji. Tak jest w istocie. Prędkość przechodzenia informacji poprzez synapsę jest niewspółmiernie mniejsza od prędkości przewodzenia wzdłuż włókien nerwowych. Jest to zrozumiałe ze wzglądu na odmienność podłoża i charakter przewodzenia.
Połączenia synaptyczne pomiędzy neuronami są obszarem aktywnej działalności integracyjnej. Dzieje się tak dlatego, że potencjały synaptyczne są zjawiskami elektrycznymi stopniowanymi, w odróżnieniu od zjawisk typu "wszystko albo nic" występujących w aksonach. Z tego powodu istnieje możliwość oddziaływania na siebie i łączenia zarówno w czasie, jak i przestrzeni. Nawet pojedyncza synapsa może być obszarem integracji poprzez sumowanie i torowanie potencjałów synaptycznych. Jeżeli weźmie się jeszcze pod uwagę, że większość neuronów w organizmie odbiera jednocześnie impulsy z synaps pobudzających i hamujących, wtedy w pełni zdać można sobie sprawę z możliwości integracji informacji nerwowej w obrębie synaps.
Do podstawowych procesów, będących wyrazem integracji na poziomie synaps, należą: sumowanie (w czasie i przestrzeni), torowanie, hamowanie presynaptyczne, hamowanie postsynaptyczne.
W niektórych synapsach każdy pre-synaptyczny potencjał czynnościowy wywołuje postsynaptyczny potencjał czynnościowy. W takim wypadku mówi się o przenoszeniu 1 : 1. Jeżeli jednak potencjały postsynaptyczne są zjawiskami podprogowymi (nie osiągającymi poziomu depolaryzacji krytycznej dla wyzwolenia potencjału czynnościowego), wtedy postsynaptyczny potencjał czynnościowy powstanie po sumowaniu w czasie lub po torowaniu pobudzających potencjałów postsynaptycz-nych lub też po obydwu tych procesach. W sumowaniu postsynaptyczne potencjały pobudzające dodają się, natomiast podczas torowania wielkość potencjałów synaptycznych rośnie wraz z powtórzeniem bodźca. Tak może dziać się w bardzo prostym przypadku pojedynczej synapsy. Kiedy neuron postsynaptyczny otrzymuje informację od większej liczby komórek, wtedy możliwości integracji stają się większe. Obok sumowania w czasie występować może sumowanie przestrzenne.
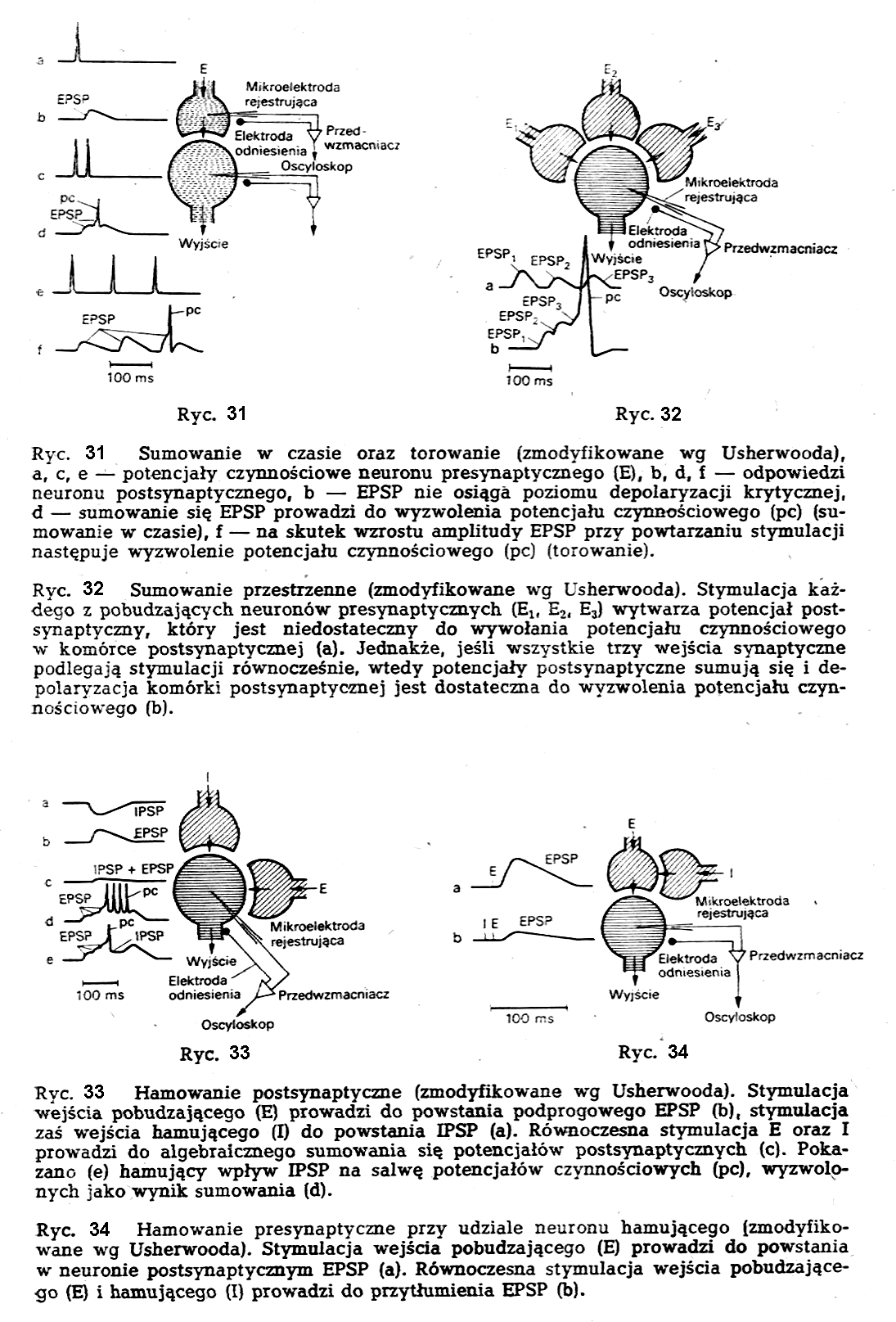
Procesy
integracyjne w obszarze synaptycznym staja się jeszcze bardziej
złożone, gdy neuron posiada wejście zarówno pobudzające, jak i
hamujące. Wtedy dochodzi do sumowania potencjałów
postsynaptycznych zarówno pobudzających, jak i hamujących.
Jest to hamowanie postsynaptyczne. Integracja synaptyczna komplikuje
się jeszcze bardziej przy zjawisku hamowania pre-synaptycznego. Ten
rodzaj hamowania związany jest z istnieniem synaps akso-aksonalnych
leżących w rejonie presy-naptycznym akso-dendrytycznej lub
akso-somatycznej synapsy pobudzającej. Intensywne badania
mikroelektro-dowe doprowadziły do ustalenia dwóch różnych
mechanizmów hamowania presynaptycznego. W pierwszym z nich
elementem presynaptycznym synapsy akso-aksonalnej jest neuron
hamujący, którego działalność doprowadza do hi-per polaryzacji
części presynaptycznej synapsy akso-dendrytycznej lub
akso--somatycznej. Hiperpolaryzacja ta uniemożliwia przewodzenie
impulsów w tym rejonie, w wyniku czego nie dochodzi do
uwolnienia mediatora, a co za tym idzie, przenoszenie zostaje
zablokowane.
Inny rodzaj hamowania presynap-tycznego może być realizowany na podłożu synapsy akso-aksonalnej o charakterze pobudzającym. W tym przypadku działalność tej synapsy doprowadza do depolaryzacji jej postsynaptycznej części. Depolaryzacja powoduje zmniejszenie amplitudy potencjałów czynnościowych biegnących w aksonie synapsy akso-dendrytycznej lub akso-somatycznej. Ilość uwalnianego mediatora zależy nie tylko od częstotliwości, ale także od amplitudy potencjałów czynnościowych dochodzących do rejonu prę-synaptycznego. Opisane zmniejszenie amplitudy potencjałów czynnościowych doprowadza więc w rezultacie do zmniejszenia ilości uwalnianego mediatora, co może spowodować zahamowanie procesu przenoszenia synaptycznego. Ten typ hamowania presynaptycznego charakteryzuje sią działaniem to-nicznym, tzn. może utrzymywać się przez diuższy okres (do 200 ms).
Opisane tu mechanizmy integracji na poziomie synaps przedstawiono i objaśniono bardziej szczegółowo na ryc. 31-34.
Biosynteza, transport wewnątrzkomórkowy, magazynowanie i uwalnianie mediatorów synaptycznych pobudzających i hamujących
W 1958 roku Paton zaproponował następujące kryteria, które winna spełniać substancja chemiczna, aby mogła być zaklasyfikowana jako mediator chemiczny:
1)Substancja ta winna być zawarta w neuronie presynaptycznym, który winien posiadać zdolność do jej syntezy.
2)Na skutek stymulacji aksonu presynaptycznego substancja ta powinna być uwalniana.
3)Podziałanie tą substancja na komórkę postsynaptyczną powinno prowadzić do efektu charakterystycznego dla normalnego przenoszenia poprzez badaną synapsę.
4)Działanie badanej substancji na komórkę postsynaptyczną powinno podlegać wpływom środków blokujących w ten sam sposób, w jaki ich wpływom podlega normalne przenosze nie synaptyczne.
5)W okolicy działania mediatora powinien znajdować się enzym zdolny do jego unieczynnienia przez rozkład.
Piąte kryterium ukazuje jedynie jeden ze sposobów unieczynnienia mediatora chemicznego. Obok rozkładu enzymatycznego mediator synaptyczny może być usuwany ze szczeliny synaptycznej przez dyfuzję lub reabsorpcję.
Cały proces działania mediatorów w obrębie synapsy można podzielić na kolejne, zazębiające się etapy: biosynteza, transport wewnątrzkomórkowy, maga--zynowanie, uwalnianie, łączenie się z receptorem błony postsynaptycznej, unieczynnianie. Synteza mediatorów synaptycznych zachodzić może w zasadzie w całym obszarze neuronu, choć zintensyfikowanie tego procesu obserwuje się przede wszystkim w zakończeniach aksortu. Obecnie nauka dysponuje już danymi dotyczącymi szczegółowych cyklów syntezy mediatorów chemicznych, w których to cyklach wiodącą role odgrywają układy enzymatyczne. W zależności od miejsca syntezy wypływa sprawa wewnątrzkomórkowego transportu mediatora. W 1968 roku opubli-wana została przez Schmitta teoria o transporcie tzw. pęcherzyków synaptycznych (zawierających mediator) od ciała neuronu wzdłuż aksonu do jego zakończeń. W procesie tym ważną rolę odgrywają neurotubule aksonalne, wzdłuż których odbywa się transport wewnątrzkomórkowy.
Miejscem magazynowania przenośnika synaptycznego są przede wszystkim pęcherzyki synaptyczne. Pęcherzyki te o średnicy 20-60 nm są częścią składowa 'kolbek synaptycznych. Te ostatnie są strukturami tworzącymi zakończenie aksonu.
Proces uwalniania mediatora synaptycznego ma kilka charakterystycznych cech. Chemiczne przenoszenie synaptyczne może dojść do skutku jedynie wtedy, gdy istnieje substancja przenośnikowa gotowa do uwolnienia do szczeliny synaptycznej w chwili, gdy błona presynaptyczna zostanie zdepolaryzowana. Pierwszą z charakterystycznych cech uwalniania mediatora synaptycznego jest uwalnianie tzw. pakietów mediatora, a więc pewnej, określonej liczby cząsteczek przenośnika (quantal re-lease). Fakt ten jest poparciem dla hipotezy pęcherzykowej, gdyż wtedy pakiet mediatora można sobie wyobrazić jako zawartość jednego pęcherzyka synaptycznego. Zakłada się np., że w jednym pęcherzyku synaptycznym synapsy chołinergicznej znajduje się ok. 1000 cząsteczek acetylocholiny.
Drugą cechą charakterystyczną uwalniania mediatora jest to, że mediator uwalnia się z zakończeń aksonu przez cały czas, niezależnie od tego, czy do zakończeń aksonu dociera impuls -czy nie. Fakt ten może być w pełni zrozumiały, jeśli się doda, że między uwalnianiem "spoczynkowym" a "czynnościowym" istnieją duże różnice ilościowe. W czasie, gdy do zakończeń aksonu nie dociera impulsacja nerwowa, z zakończeń tych uwalniają się spontanicznie małe ilości mediatora. Istnieje przypuszczenie, że zjawisko to można przypisać faktowi, iż pęcherzyki synaptyczne znajdując się w ciągłym ruchu, w pewnych momentach stykają się z "obszarem uwalniania" (tzw. obszar aktywny) i uwalniają swą zawartość do szczeliny synaptycznej. Depolaryzacja biony presynaptycznej przez potencjał czynnościowy prowadzi do bardzo znacznego zwiększenia ,,obszaru uwalniania", wzrasta wówczas możliwość zetknięcia się pęcherzyków synaptycznych z takim obszarem, W ten sposób do szczeliny synaptycznej uwolniona zostaje równocześnie duża liczba "pakietów".
Trzecią wreszcie charakterystyczną cechą procesu uwalniania mediatora jest rola jonów wapniowych w tym procesie. Okazało się, że pod nieobecność jonów Ca++ w płynie zewnątrzkomór-kowym depolaryzacja aksonu przez potencjał czynnościowy nie doprowadza do zwiększonego uwalniania mediatora. W takich warunkach spontaniczne uwalnianie małych ilości mediatora nie zostaje zatrzymane. Można z tego wysunąć wniosek, że Ca++ jest niezbędny w procesie uruchamiania dodatkowych "obszarów uwalniania" w bio-nie presynaptycznej. Mechanizmem spustowym aktywacji wapnia (wzrost przepuszczalności błony presynaptycznej dla tego jonu) jest depolaryzacja obszaru presynaptycznego. Podobne efekty do spowodowanych niedoborem lub nieobecnością Ca++ obserwuje się, stosując na obszar synaptyczny zwiększone stężenie Mg++. Hamujące działanie Mg++ na przenoszenie synaptyczne tłumaczone jest kompetytywnym działaniem tych jonów na obszary błony presynaptycznej "zarezerwowane" dia jonów wapniowych.
Wydzielony z części presynaptycznej mediator dyfunduje poprzez szczelinę synaptyczną w kierunku błony postsynaptycznej. Następuje następny etap działalności mediatora: łączenie się z receptorem błony postsynaptycznej. Receptory błonowe są zdefiniowanymi chemicznie obszarami dużych cząsteczek, które łączą się z mediatorem na zasadzie "chemicznego dopełnienia". Jako wynik połączenia się mediatora z receptorem błonowym powstają zmiany przepuszczalności błony postsynaptycznej dla różnych jonów, a w dalszej kolejności zjawiska bioelektryczne, nazywane ogólnie zjawiskami postsynaptycznymi.
Spontaniczne uwalnianie mediatora powoduje powstanie w błonie postsynaptycznej tzw. miniaturowych potencjałów postsynaptycznych. Potencjały te, o amplitudzie ok. 0,7 mV, stwierdzono i opisano najpierw na preparatach nerwowo-miąśniowych. Okazało się jednak, że i synapsy nerwowo-nerwowe są miejscem powstawania miniaturowych potencjałów postsynaptycznych. Potencjały te uważa się za wynik uwalniania pojedynczych pakietów mediatora.
Zwiększone uwalnianie mediatora na skutek depolaryzacji błony presynap-tycznej prowadzi do powstania w części postsynaptycznej typowych postsynaptycznych potencjałów pobudzających (EPSP), lub też postsynaptycznych potencjałów hamujących (IPSP), w zależności od charakteru synapsy. Pobudzające potencjały postsynaptyczne dają początek potencjałom czynnościowym.
W 1970 r. dwaj badacze brytyjscy Katz i Miledi opisali jeszcze jedno postsynaptyczne zjawisko bioelektryczne. Pracując na preparacie nerwowo-mięśniowym żaby, wyodrębnili oni w błonie postsynaptycznej potencjały o amplitudzie 0,3 V, które według ich hipotezy mają być odzwierciedleniem połączenia się jednej cząsteczki mediatora z receptorem cholinergicznym. Potencjały te nazwane zostały szumem acetylocholinowym.
Ostatnim wreszcie etapem losów mediatora w obrębie synapsy jest jego unieczynnienie. Jednym ze sposobów unieczynnienia mediatora jest jego rozkład za pomocą enzymu. Taki los spotyka np. a c ety locho linę, która jest rozkładana przez acetylocholine-sterazą na cbołinę i kwas octowy. Istnieją wszakże inne, bardziej powszechne, sposoby unieczynniania mediatorów synaptycznych. Do nich należy wychwyt mediatora przez elementy presynaptyczne, czy też glej, jak to ma miejsce np. w przypadku amin katecholo-wych, kwasu gammaa min oma słów ego, czy też glicyny.
Mimo intensywnych badań w zakresie identyfikacji mediatorów synaptycznych, w wielu obszarach układu nerwowego nie udało się ustalić, jakie substancje pełnią rolę przenoszącą. Spośród mediatorów pobudzających najbardziej zasadniczą rolę spełnia acety-locholina (ACh). Istnieje cały szereg związków, którym poza acetylocholiną przypisywane są właściwości chemicznych mediatorów stanów pobudzających. Spośród mediatorów hamujących, wywołujących hiperpolaryzację neuronu postsynaptycznego, na pierwsze miejsce wysuwa się kwas gamma-ami-nomasłowy (GABA). Znaczna część synaps w ośrodkowym układzie nerwowym gromadzi ten związek.
W okresie kilkudziesięciu lat ogólnie przyjęty był pogląd, że jeden neuron może syntetyzować i uwalniać ze swych zakończeń jedynie jeden mediator chemiczny. Jest to tzw. prawo Dale'ar który wraz z Loewim w 1936 r. otrzymał nagrodę Nobla za prace z zakresu chemicznych mediatorów synaptycznych.
Najnowsze badania wykazują jednak, że występują neurony syntetyzujące dwa mediatory synaptyczne.
KODOWANIE INFORMACJI PRZEZ KOMÓRKI NERWOWE
Bodźce działające na układ nerwowy zawierają informacje pochodzące zarówno z wnętrza organizmu, jak i z jego otoczenia. Bodźce te reprezentują rozmaite formy energii. Prawidłowa działalność układu nerwowego, a co za tym idzie, całego organizmu, wymaga w związku z poruszonym tu problemem obecności dwóch procesów. Pierwszym jest proces przetworzenia energii związanej z danym bodźcem na impulsację elektryczną, a więc na zjawiska bioelektryczne. Drugim zaś procesem jest kodowanie odebranej informacji w obrąbie układu nerwowego.
Wydawałoby się, że jednym z najprostszych sposobów kodowania informacji w obrębie układu nerwowego mógłby być kod amplitudy. Układ nerwowy nie korzysta jednak z tego sposobu informacji (por. prawo "wszystko albo nic"). Informacja biegnąca w układzie nerwowym zakodowana jest za pomocą kodu częstości. W tym rodzaju kodowania miarą przenoszonego parametru informacji jest gęstość impulsów. W odróżnieniu od kodu interwałowego w kodzie częstości nie chodzi o przerwy miedzy impulsami, lecz o średnią częstość impulsów w określonym przedziale czasu. Jak widać z założeń tego sposobu kodowania, zmiana amplitudy potencjałów, lub też "zgubienie" pojedynczych impulsów nie może wpłynąć w istotny sposób na treść zakodowanej informacji. Według współczesnych poglądów, pojemność informacyjna (zdolność przenoszenia) danego kanału komunikacyjnego rozpatrywana jest w kategoriach szybkości przekazywania informacji. Według definicji pojemność informacyjna kanału to maksymalna ilość informacji przesyłanych w jednostce czasu (bit/s).

Spośród
koncepcji dotyczących określania pojemności informacji w kanałach
nerwowych na uwagę zasługuje następująca, wyrażona wzorem:
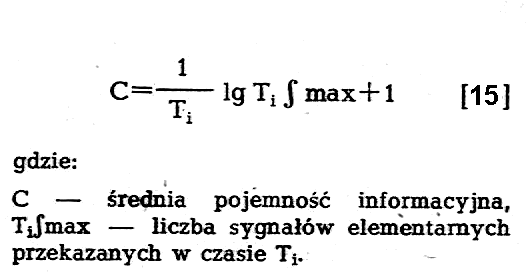
Wzór
ten bierze pod uwagą tzw. czynność spontaniczną.
Impulsacja spontaniczna występuje w wielu kanałach nerwowych przy braku sygnału (bodźca). Bardzo często impulsacja ta z punktu widzenia przesyłania sygnałów traktowana jest jako zakłócenie. W istocie jednak odgrywa ona ważną rolę przy odbiorze słabych sygnałów. Jeśli przyjmiemy, że częstość impulsacji we włóknach nerwowych rośnie wprost proporcjonalnie do natężenia bodźca, według funkcji zbliżonej do logarytmicznej, wtedy iatwo sobie wyobrazić, że dla bardzo słabych bodźców częstotliwość impulsacji zbliżałaby się do zera. Mogłoby to wpływać w istotny sposób na opóźnienie odpowiedzi na słabe bodźce. Istnienie impulsacji spontanicznej i traktowanie informacji jako czynnika zmieniającego te impulsację rozwiązuje w większości przypadków problem informacji o bardzo słabych bodźcach.
Wiele powyższych koncepcji opracowano na podstawie założeń teoretycznych wynikających z modelowania czynności neuronu w ramach prac wchodzących w zakres Moniki. Z drugiej strony istnieje szereg danych doświadczalnych potwierdzających powyższe założenia.
Na ryc. 35 do 37 przedstawiono przykłady kodowania informacji w neuronach.
Powyższy artukuł nie wyczermuje całości zagadnienia. Jest tylko przybliżeniem i zasygnalizowaniem złożoności tematu.